

Regulation of chromosome stability by the histone H2A variant Htz1, the Swr1 chromatin remodeling complex, and the histone acetyltransferase NuA4
- PMID: 15353583
- PMCID: PMC518788
- DOI: 10.1073/pnas.0405753101
Regulation of chromosome stability by the histone H2A variant Htz1, the Swr1 chromatin remodeling complex, and the histone acetyltransferase NuA4
Erratum in
- Proc Natl Acad Sci U S A. 2006 Apr 18;103(16):6410
Abstract
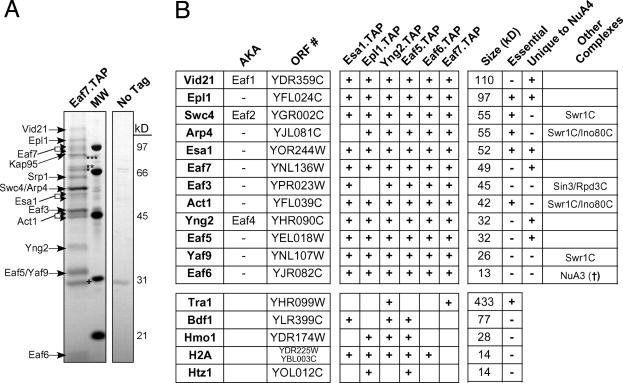
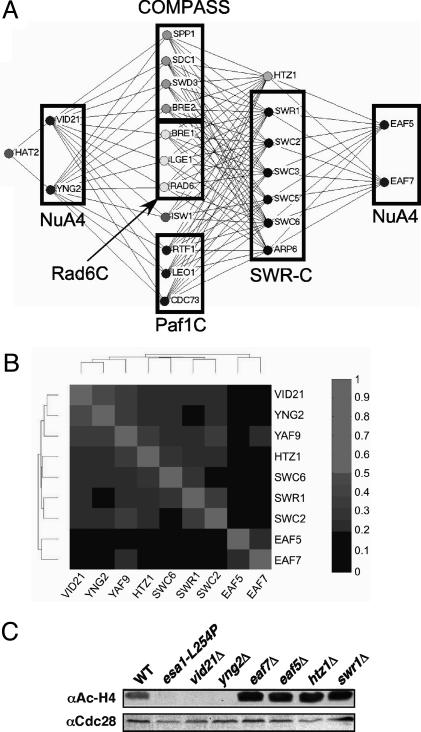
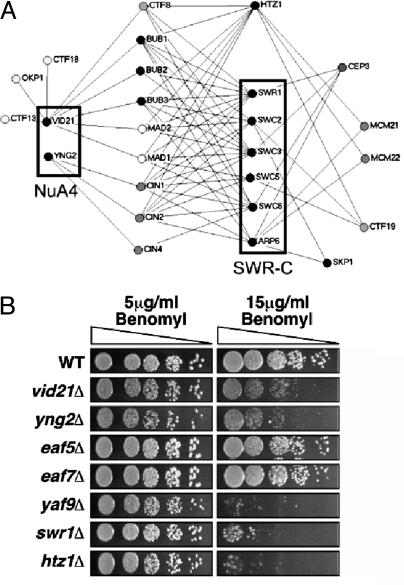
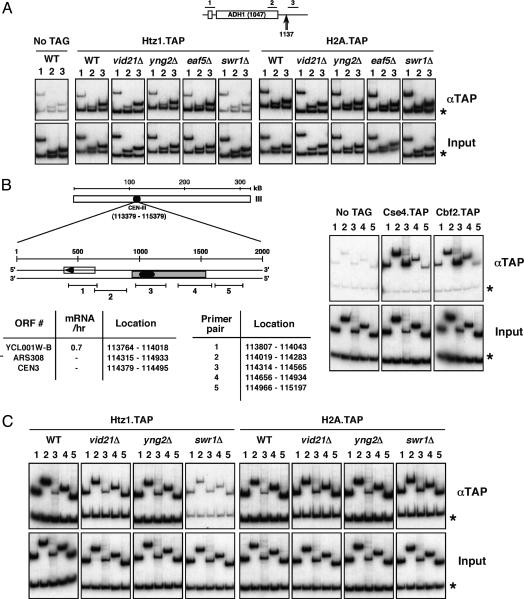
NuA4, the only essential histone acetyltransferase complex in Saccharomyces cerevisiae, acetylates the N-terminal tails of histones H4 and H2A. Affinity purification of NuA4 revealed the presence of three previously undescribed subunits, Vid21/Eaf1/Ydr359c, Swc4/Eaf2/Ygr002c, and Eaf7/Ynl136w. Experimental analyses revealed at least two functionally distinct sets of polypeptides in NuA4: (i) Vid21 and Yng2, and (ii) Eaf5 and Eaf7. Vid21 and Yng2 are required for bulk histone H4 acetylation and are functionally linked to the histone H2A variant Htz1 and the Swr1 ATPase complex (SWR-C) that assembles Htz1 into chromatin, whereas Eaf5 and Eaf7 have a different, as yet undefined, role. Mutations in Htz1, the SWR-C, and NuA4 cause defects in chromosome segregation that are consistent with genetic interactions we have observed between the genes encoding these proteins and genes encoding kinetochore components. Because SWR-C-dependent recruitment of Htz1 occurs in both transcribed and centromeric regions, a NuA4/SWR-C/Htz1 pathway may regulate both transcription and centromere function in S. cerevisiae.
Figures
Similar articles
-
The Saccharomyces cerevisiae histone H2A variant Htz1 is acetylated by NuA4.Genes Dev. 2006 Mar 15;20(6):660-5. doi: 10.1101/gad.1388106. Genes Dev. 2006. PMID: 16543219 Free PMC article.
-
NuA4-dependent acetylation of nucleosomal histones H4 and H2A directly stimulates incorporation of H2A.Z by the SWR1 complex.J Biol Chem. 2010 May 21;285(21):15966-77. doi: 10.1074/jbc.M110.117069. Epub 2010 Mar 23. J Biol Chem. 2010. PMID: 20332092 Free PMC article.
-
The Yaf9 component of the SWR1 and NuA4 complexes is required for proper gene expression, histone H4 acetylation, and Htz1 replacement near telomeres.Mol Cell Biol. 2004 Nov;24(21):9424-36. doi: 10.1128/MCB.24.21.9424-9436.2004. Mol Cell Biol. 2004. PMID: 15485911 Free PMC article.
-
NuA4 and SWR1-C: two chromatin-modifying complexes with overlapping functions and components.Biochem Cell Biol. 2009 Oct;87(5):799-815. doi: 10.1139/O09-062. Biochem Cell Biol. 2009. PMID: 19898529 Review.
-
Variation on a theme: Evolutionary strategies for H2A.Z exchange by SWR1-type remodelers.Curr Opin Cell Biol. 2021 Jun;70:1-9. doi: 10.1016/j.ceb.2020.10.014. Epub 2020 Nov 17. Curr Opin Cell Biol. 2021. PMID: 33217681 Review.
Cited by
-
Aberrant Meiotic Prophase I Leads to Genic Male Sterility in the Novel TE5A Mutant of Brassica napus.Sci Rep. 2016 Sep 27;6:33955. doi: 10.1038/srep33955. Sci Rep. 2016. PMID: 27670217 Free PMC article.
-
Systematic yeast synthetic lethal and synthetic dosage lethal screens identify genes required for chromosome segregation.Proc Natl Acad Sci U S A. 2005 Sep 27;102(39):13956-61. doi: 10.1073/pnas.0503504102. Epub 2005 Sep 19. Proc Natl Acad Sci U S A. 2005. PMID: 16172405 Free PMC article.
-
The Saccharomyces cerevisiae histone H2A variant Htz1 is acetylated by NuA4.Genes Dev. 2006 Mar 15;20(6):660-5. doi: 10.1101/gad.1388106. Genes Dev. 2006. PMID: 16543219 Free PMC article.
-
Targeting of the SUN protein Mps3 to the inner nuclear membrane by the histone variant H2A.Z.J Cell Biol. 2011 May 2;193(3):489-507. doi: 10.1083/jcb.201011017. Epub 2011 Apr 25. J Cell Biol. 2011. PMID: 21518795 Free PMC article.
-
Histone variant H2A.Z is needed for efficient transcription-coupled NER and genome integrity in UV challenged yeast cells.PLoS Genet. 2024 Sep 10;20(9):e1011300. doi: 10.1371/journal.pgen.1011300. eCollection 2024 Sep. PLoS Genet. 2024. PMID: 39255275 Free PMC article.
References
-
- Lusser, A. & Kadonaga, J. T. (2003) BioEssays 25, 1192–1200. - PubMed
-
- Strahl, B. D. & Allis, C. D. (2000) Nature 403, 41–45. - PubMed
-
- Jenuwein, T. & Allis, C. D. (2001) Science 293, 1074–1080. - PubMed
-
- Malik, H. S. & Henikoff, S. (2003) Nat. Struct. Biol. 10, 882–891. - PubMed
-
- Krogan, N. J., Keogh, M.-C., Datta, N., Sawa, C., Ryan, O. W., Ding, H., Haw, R. A., Pootoolal, J., Tong, A., Canadien, V., et al. (2003) Mol. Cell 12, 1565–1576. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
