

Keeping sensory cells and evolving neurons to connect them to the brain: molecular conservation and novelties in vertebrate ear development
- PMID: 15353909
- PMCID: PMC1242196
- DOI: 10.1159/000079746
Keeping sensory cells and evolving neurons to connect them to the brain: molecular conservation and novelties in vertebrate ear development
Abstract
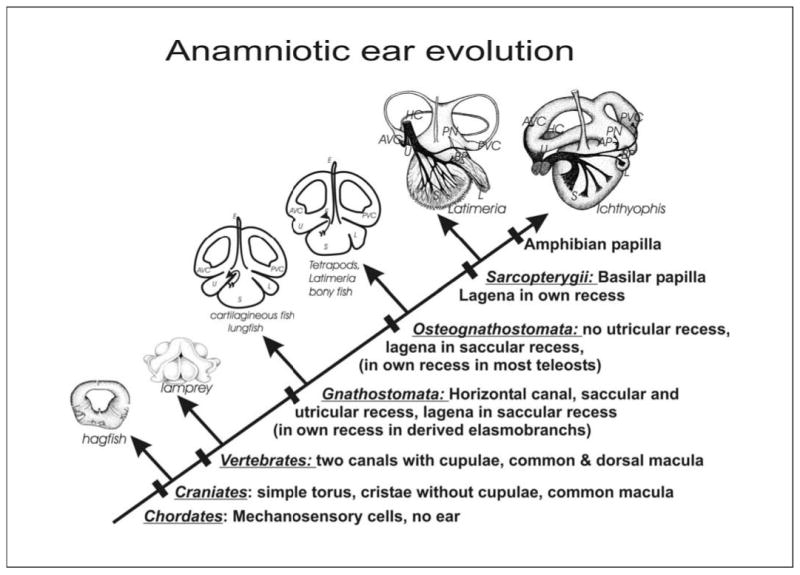
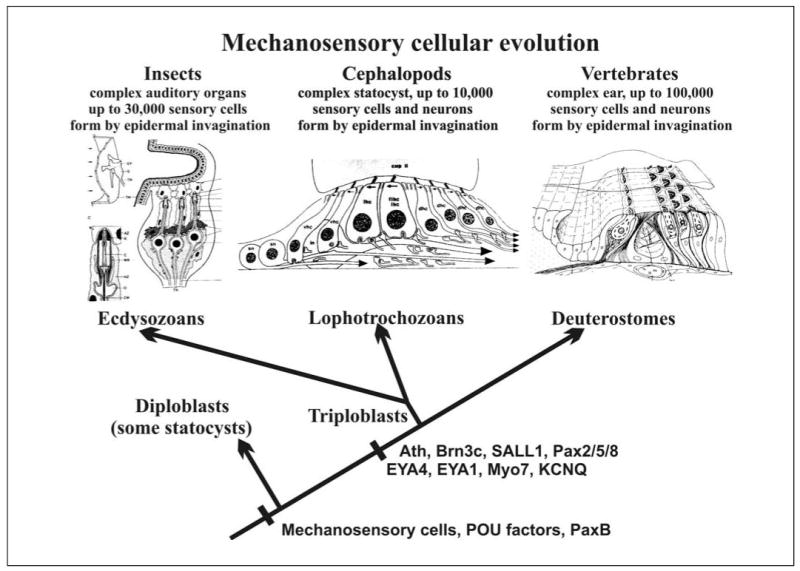
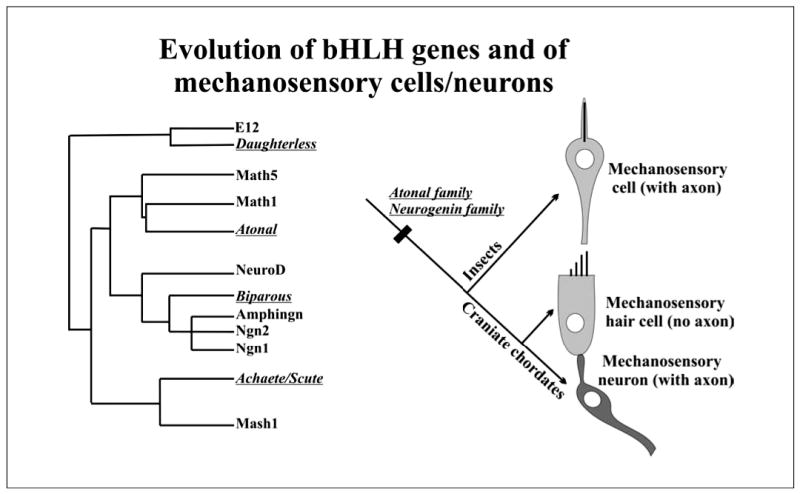
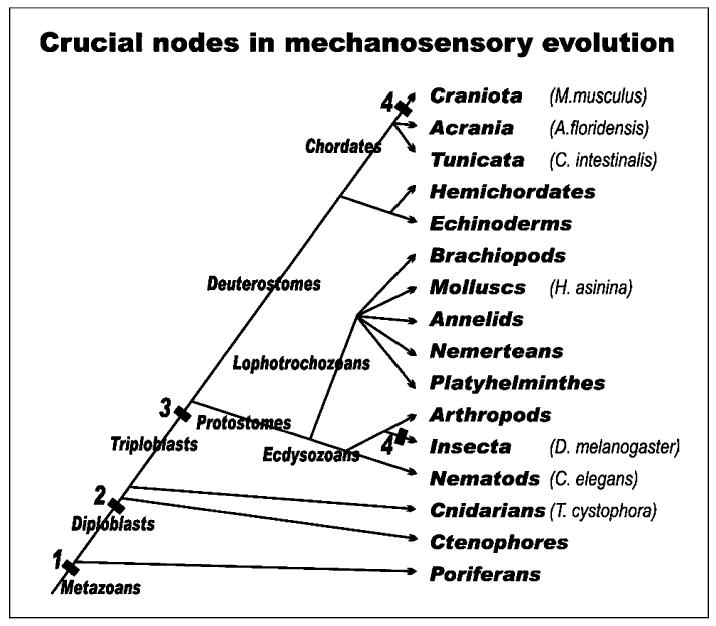
The evolution of the mechanosensory cellular module and the molecular details that regulate its development has included morphological modifications of these cells as well as the formation of larger assemblies of mechanosensory cell aggregates among metazoans. This has resulted in a wide diversity of mechanosensory organs. The wide morphological diversity of organs, including the associated morphological modifications of the mechanosensory cells, suggests parallel evolution of these modules and their associated organs. This morphological diversity is in stark contrast to the molecular conservation of developmental modules across phyla. These molecular data suggest that the evolution of mechanosensory transduction might have preceded that of distinct cellular differentiation. However, once a molecular network governing development of specialized cells involved in mechanosensory transduction evolved, that molecular network was preserved across phyla. Present data suggest that at least the common ancestor of triploblastic organisms, perhaps even the common diploblastic ancestor of bilaterian metazoans, had molecular and cellular specializations for mechanosensation. It is argued that the evolution of multicellular organs dedicated to specific aspects of mechanosensation, such as gravity and sound perception, are evolutionary transformations that build on this conserved molecular network for cellular specialization, but reflect distinct morphological solutions. We propose that the sensory neurons, connecting the craniate ear with the brain, are a derived feature of craniates, and possibly chordates, that came about through diversification of the lineage forming mechanosensory cells during development. This evolutionarily late event suggests a heterochronic shift, so that sensory neurons develop in mammals prior to mechanosensory hair cells. However, sensory neuron development is connected to hair cell development, likely in a clonal relationship. The theme of cellular conservation is reiterated in two examples of chordate otic diversification: the evolution of the horizontal canal system and the evolution of the basilar papilla/cochlea. It is suggested that here again, cellular multiplication and formation of a special epithelium predates the functional transformation to an 'organ' system for horizontal angular acceleration and sound pressure reception, respectively. Overall, evolution of the vertebrate ear needs to be understood as an interplay between and utilization of two gene networks or modules. One is at the level of the molecularly and developmentally conserved mechanosensory cellular module. The other is an increased complexity in the morphology of both adult mechanosensory cells and organs by the addition of end-stage and novel features and associated gene networks to detect specific aspects of mechanosensory stimuli.
Figures
Similar articles
-
Molecular evolution of the vertebrate mechanosensory cell and ear.Int J Dev Biol. 2007;51(6-7):663-78. doi: 10.1387/ijdb.072367bf. Int J Dev Biol. 2007. PMID: 17891725 Free PMC article. Review.
-
Cells, molecules and morphogenesis: the making of the vertebrate ear.Brain Res. 2006 May 26;1091(1):151-71. doi: 10.1016/j.brainres.2006.02.078. Epub 2006 Apr 27. Brain Res. 2006. PMID: 16643865 Free PMC article. Review.
-
Evolution of sound and balance perception: innovations that aggregate single hair cells into the ear and transform a gravistatic sensor into the organ of corti.Anat Rec (Hoboken). 2012 Nov;295(11):1760-74. doi: 10.1002/ar.22573. Epub 2012 Oct 8. Anat Rec (Hoboken). 2012. PMID: 23044863 Review.
-
Developmental evolutionary biology of the vertebrate ear: conserving mechanoelectric transduction and developmental pathways in diverging morphologies.Neuroreport. 2000 Nov 27;11(17):R35-44. doi: 10.1097/00001756-200011270-00013. Neuroreport. 2000. PMID: 11117521 Review.
-
Evolution of vertebrate mechanosensory hair cells and inner ears: toward identifying stimuli that select mutation driven altered morphologies.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2014 Jan;200(1):5-18. doi: 10.1007/s00359-013-0865-z. Epub 2013 Nov 27. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2014. PMID: 24281353 Free PMC article. Review.
Cited by
-
Hedgehog signaling governs the development of otic sensory epithelium and its associated innervation in zebrafish.J Neurosci. 2010 Mar 10;30(10):3612-23. doi: 10.1523/JNEUROSCI.5109-09.2010. J Neurosci. 2010. PMID: 20219995 Free PMC article.
-
Smaller inner ear sensory epithelia in Neurog 1 null mice are related to earlier hair cell cycle exit.Dev Dyn. 2005 Nov;234(3):633-50. doi: 10.1002/dvdy.20551. Dev Dyn. 2005. PMID: 16145671 Free PMC article.
-
Molecular evolution of the vertebrate mechanosensory cell and ear.Int J Dev Biol. 2007;51(6-7):663-78. doi: 10.1387/ijdb.072367bf. Int J Dev Biol. 2007. PMID: 17891725 Free PMC article. Review.
-
Development and evolution of the vestibular sensory apparatus of the mammalian ear.J Vestib Res. 2005;15(5-6):225-41. J Vestib Res. 2005. PMID: 16614470 Free PMC article. Review.
-
Cells, molecules and morphogenesis: the making of the vertebrate ear.Brain Res. 2006 May 26;1091(1):151-71. doi: 10.1016/j.brainres.2006.02.078. Epub 2006 Apr 27. Brain Res. 2006. PMID: 16643865 Free PMC article. Review.
References
-
- Alvarez Y, Alonso MT, Vendrell V, Zelarayan LC, Chamero P, Theil T, Bosl MR, Kato S, Maco-nochie M, Riethmacher D, Schimmang T. Requirements for FGF3 and FGF10 during inner ear formation. Development. 2003;130:6329–633. - PubMed
-
- Andermann P, Ungos J, Raible DW. Neurogenin1 defines zebrafish cranial sensory ganglia precursors. Dev Biol. 2002;251:45–58. - PubMed
-
- Arkett SA, Mackie GO, Meech RW. Hair cell mechanoreception in a jellyfish Aglantha digitale. J Exp Biol. 1988;135:329–342.
-
- Ben-Arie N, Hassan BA, Bermingham NA, Malicki DM, Armstrong D, Matzuk M, Bellen HJ, Zoghbi HY. Functional conservation of atonal and Math1 in the CNS and PNS. Development. 2000;127:1039–1048. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
