

The management of extracellular ice by petioles of frost-resistant herbaceous plants
- PMID: 15355865
- PMCID: PMC4242212
- DOI: 10.1093/aob/mch191
The management of extracellular ice by petioles of frost-resistant herbaceous plants
Abstract
Background and aims: Some frost-tolerant herbaceous plants droop and wilt during frost events and recover turgor and posture on thawing. It has long been known that when plant tissues freeze, extracellular ice forms. Distributions of ice and water in frost-frozen and recovered petioles of Trifolium repens and Escholschzia californica were visualized.
Methods: Petioles of intact plants were cryo-fixed, planed to smooth transverse faces, and examined in a cryo-SEM.
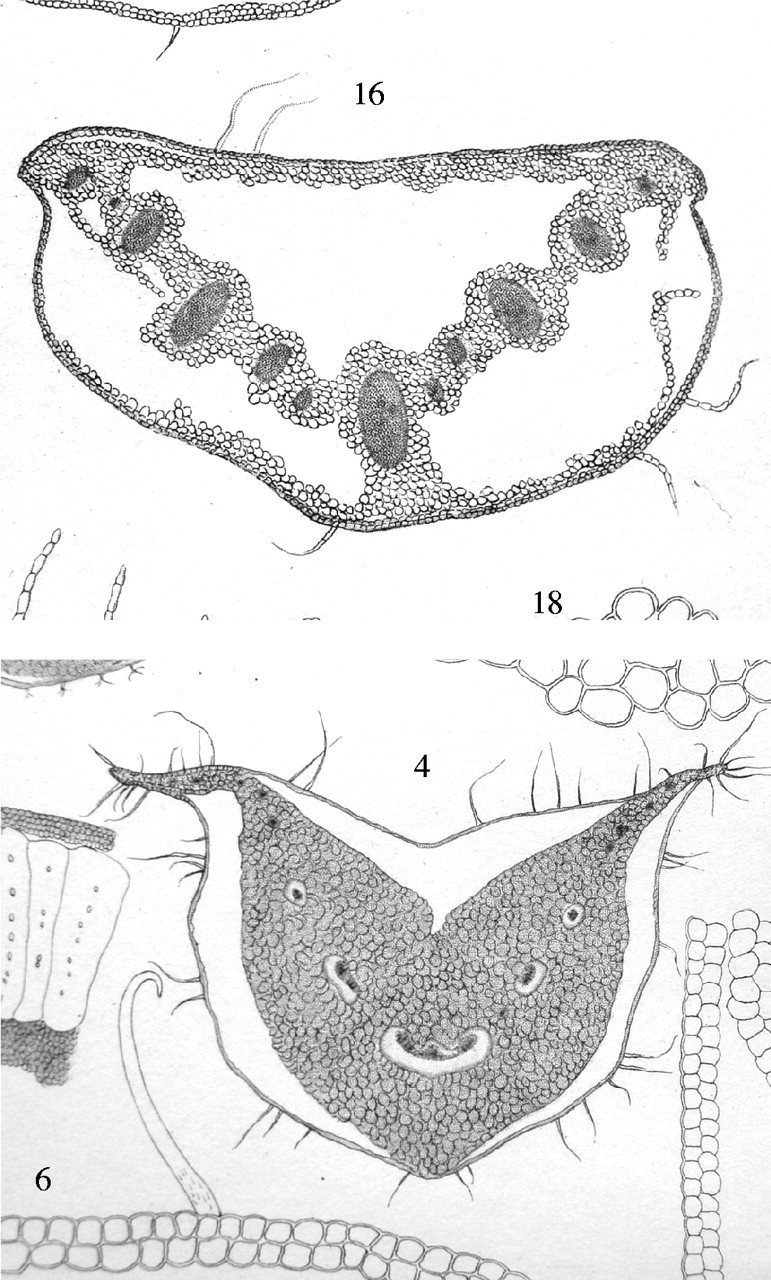
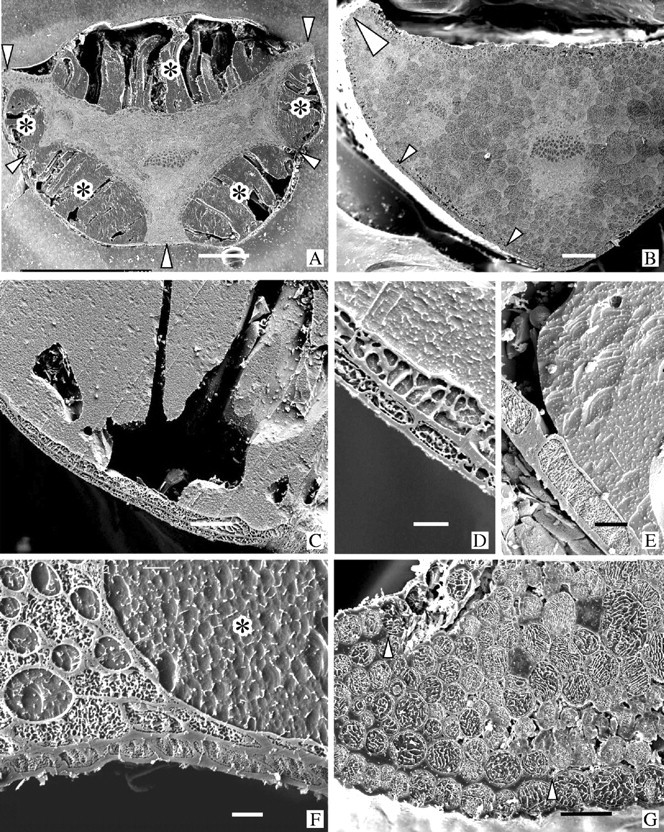
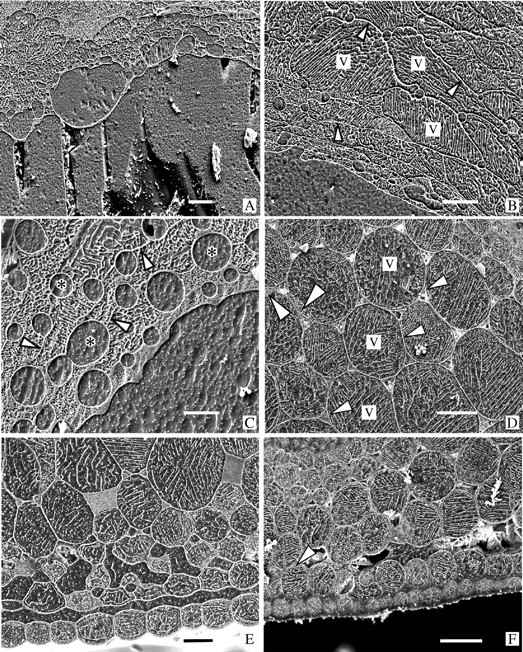
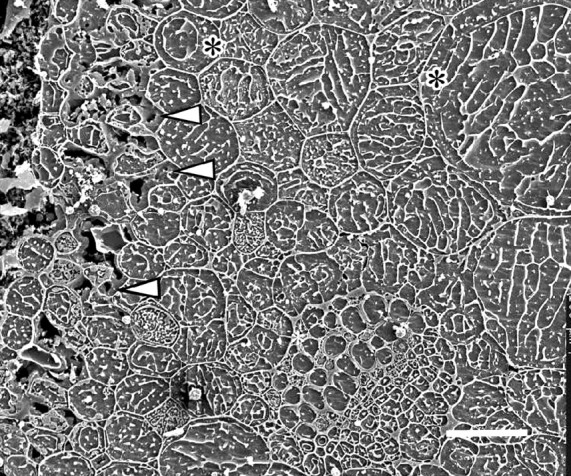
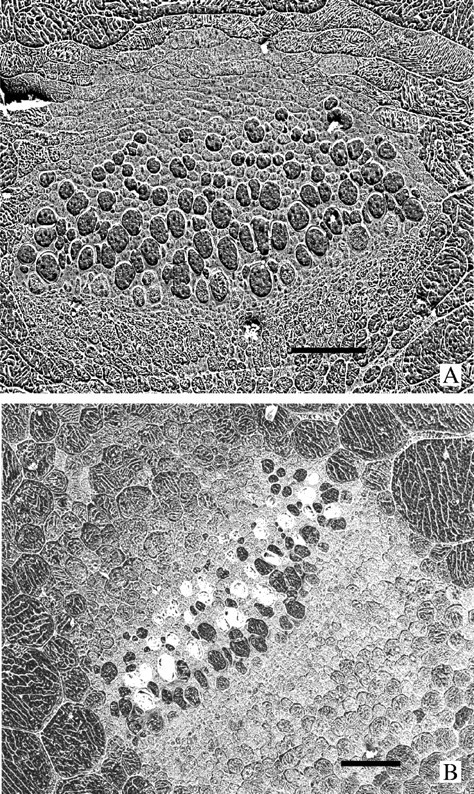
Key results: With frost-freezing, parenchyma tissues shrank to approx. one-third of their natural volume with marked cytorrhysis of the cells, and massive blocks of extracellular icicles grew under the epidermis (poppy) or epidermis and subepidermis (clover), leaving these layers intact but widely separated from the parenchyma except at specially structured anchorages overlying vascular bundles. On thawing, the extracellular ice was reabsorbed by the expanding parenchyma, and surface tissues again contacted the internal tissues at weak junctions (termed faults). These movements of water into and from the fault zones occurred repeatedly at each frost/thaw event, and are interpreted to explain the turgor changes that led to wilting and recovery. Ice accumulations at tri-cellular junctions with intercellular spaces distended these spaces into large cylinders, especially large in clover. Xylem vessels of frozen petioles were nearly all free of gas; in thawed petioles up to 20 % of vessels were gas-filled.
Conclusions: The occurrence of faults and anchorages may be expected to be widespread in frost-tolerant herbaceous plants, as a strategy accommodating extracellular ice deposits which prevent intracellular freezing and consequent membrane disruption, as well as preventing gross structural damage to the organs. The developmental processes that lead to this differentiation of separation of sheets of cells firmly cemented at determined regions at their edges, and their physiological consequences, will repay detailed investigation.
Figures

References
-
- Ashworth EN, Echlin P, Pearce RS, Hayes TL. 1988. Ice formation and tissue response in apple twigs. Plant, Cell and Environment 11: 703–710.
-
- Ashworth EN, Davis GA, Wisniewski ME. 1989. The formation and distribution of ice within dormant and deacclimated peach flower buds. Plant, Cell and Environment 12: 521–528.
-
- Ball MC, Canny MJ, Huang CX, Heady R. 2004. Structural changes in acclimated and unacclimated Eucalyptus leaves during freezing and thawing. Functional Plant Biology 31: 29–40. - PubMed
-
- Ball MC, Wolfe J, Canny M, Hofmann M, Nicotra AB, Hughes D. 2002. Space and time dependence of temperature and freezing in evergreen leaves. Functional Plant Biology 29: 1259–1272. - PubMed
