

Chronic ethanol induces synaptic but not extrasynaptic targeting of NMDA receptors
- PMID: 15356198
- PMCID: PMC6729936
- DOI: 10.1523/JNEUROSCI.1902-04.2004
Chronic ethanol induces synaptic but not extrasynaptic targeting of NMDA receptors
Abstract
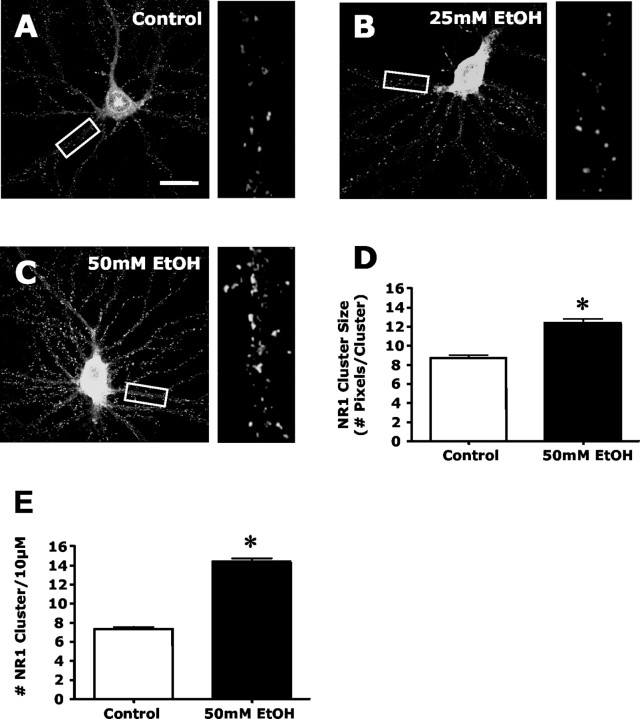
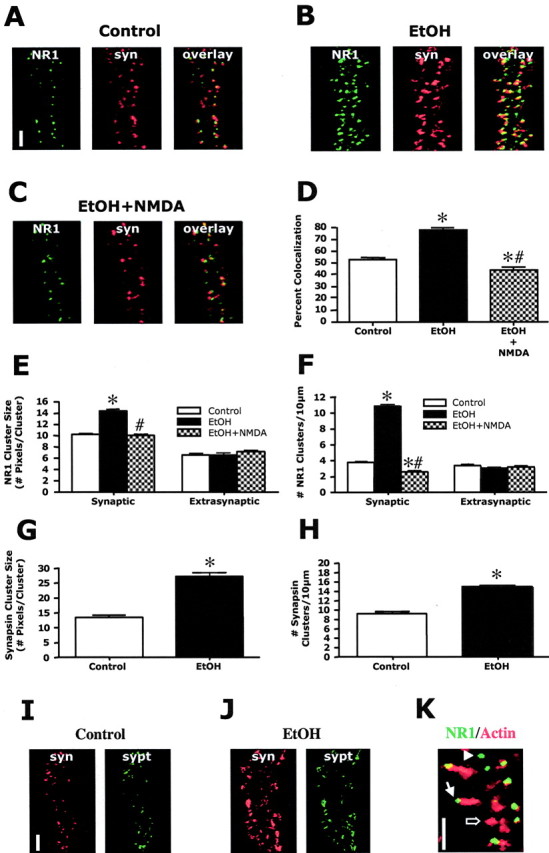
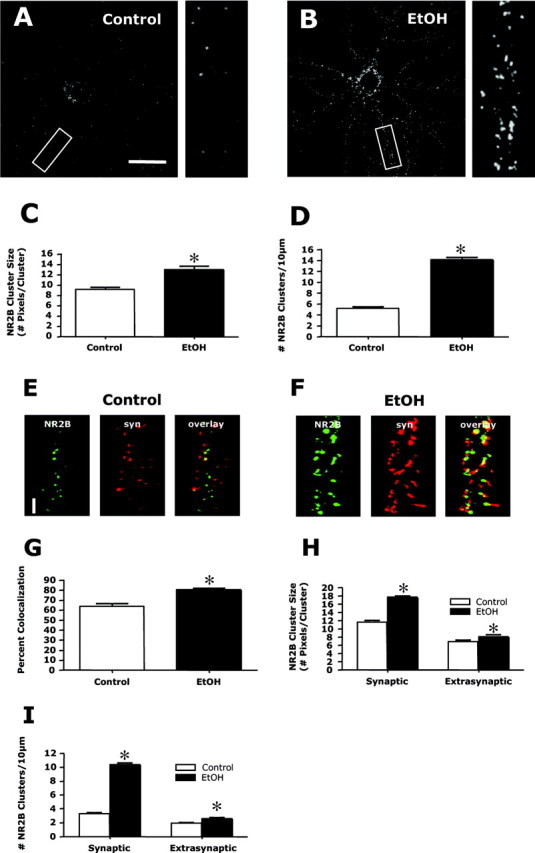
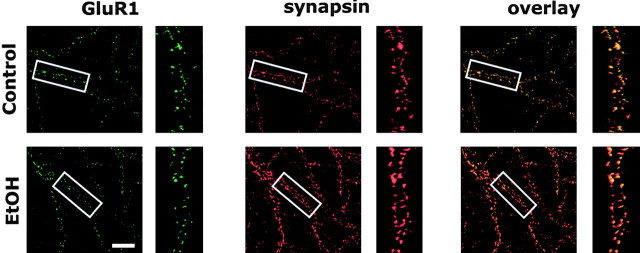
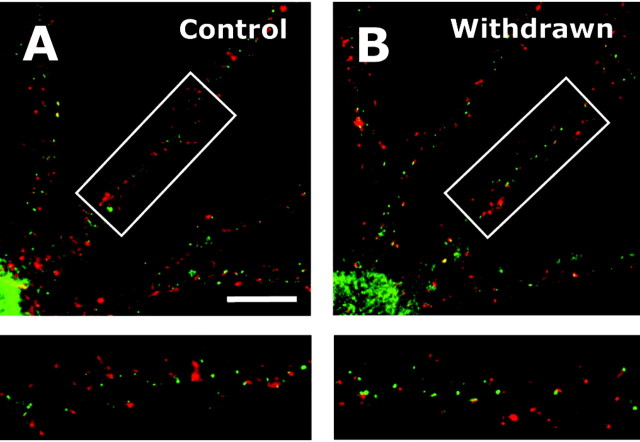
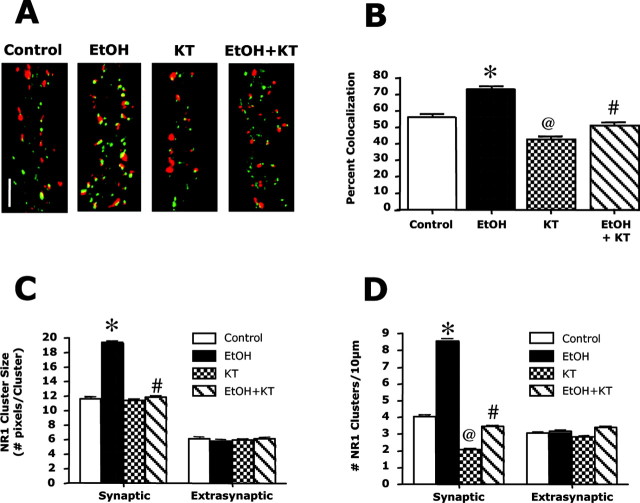
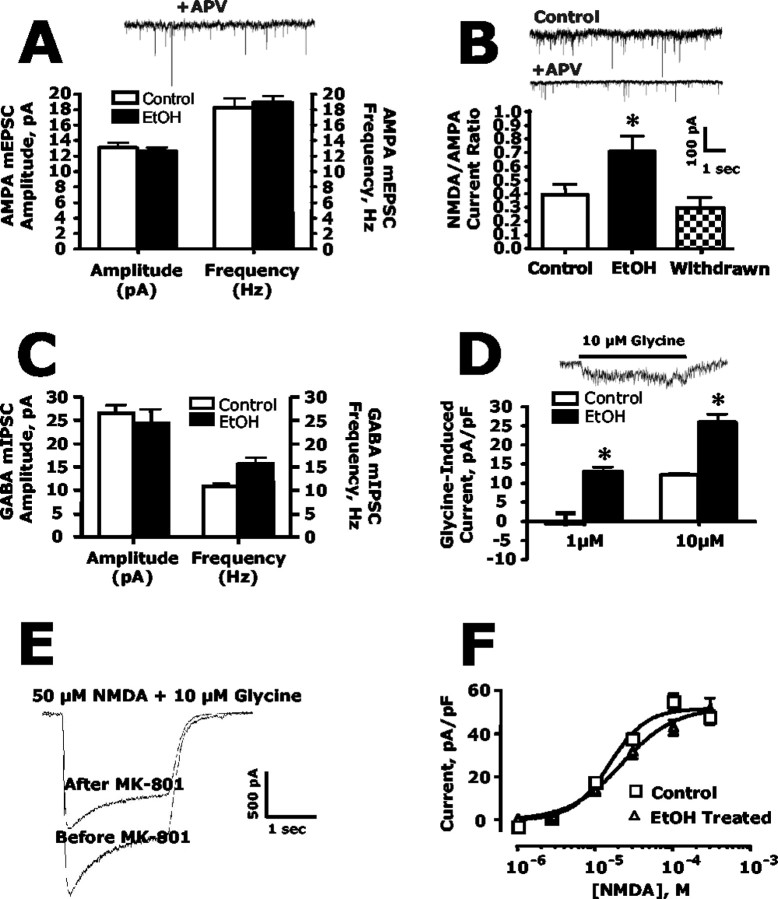
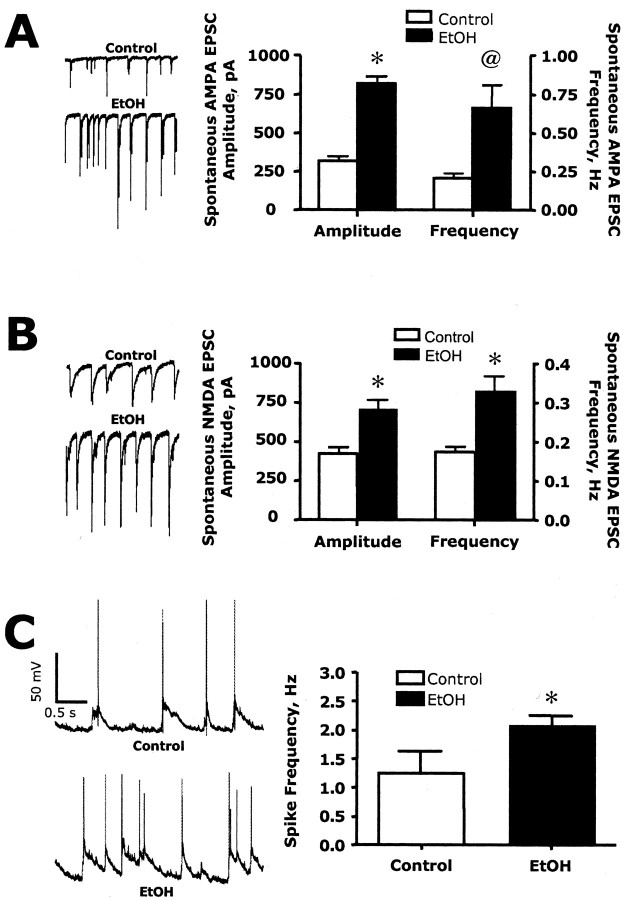
The development of ethanol tolerance and dependence reflects neuroadaptive changes in response to continuous depression in synaptic activity. The present study used confocal imaging and electrophysiology procedures to assess the effects of prolonged ethanol exposure on NMDA receptor trafficking in cultures of hippocampal neurons. Neurons exposed to 50 mm ethanol for 4 d showed an increase in the colocalization of NMDA receptor type 1 (NR1) clusters with the presynaptic marker protein synapsin. This was accompanied by significant increases in the size and density of these synapsin-associated clusters with no change observed in nonsynapsin-associated NR1 clusters. Similar effects were observed with NR2B clustering after chronic ethanol exposure. The increase in synaptic NMDA receptor clustering was prevented by addition of a protein kinase A inhibitor or by coexposure to a low concentration of NMDA and was reversed when ethanol was removed from the cultures. No changes were observed in the synaptic content, cluster size, or density of AMPA receptors after ethanol exposure. Electrophysiological measurements on ethanol-treated neurons revealed a similar enhancement in synaptic NMDA currents with no change in AMPA-mediated events. After isolation of extrasynaptic NMDA receptors by MK801 (+)-5-methyl-10,11-dihydro-5H-dibenzo [a,d] cyclohepten-5,10-imine maleate (/) trapping, whole-cell responses to NMDA were not different between control and ethanol-treated neurons These observations demonstrate that neuroadaptive changes in NMDA receptors in response to prolonged ethanol exposure occur through activity-dependent processes that regulate their synaptic targeting and localization.
Figures
Similar articles
-
Aberrant synaptic activation of N-methyl-D-aspartate receptors underlies ethanol withdrawal hyperexcitability.J Pharmacol Exp Ther. 2007 Apr;321(1):60-72. doi: 10.1124/jpet.106.111419. Epub 2007 Jan 17. J Pharmacol Exp Ther. 2007. PMID: 17229881
-
Long-term regulation of N-methyl-D-aspartate receptor subunits and associated synaptic proteins following hippocampal synaptic plasticity.Neuroscience. 2003;118(4):1003-13. doi: 10.1016/s0306-4522(03)00028-9. Neuroscience. 2003. PMID: 12732245
-
Hepatocyte growth factor improves synaptic localization of the NMDA receptor and intracellular signaling after excitotoxic injury in cultured hippocampal neurons.Exp Neurol. 2008 Mar;210(1):83-94. doi: 10.1016/j.expneurol.2007.10.001. Epub 2007 Oct 13. Exp Neurol. 2008. PMID: 18001712
-
The NR2B subtype of NMDA receptor: a potential target for the treatment of alcohol dependence.Curr Drug Targets CNS Neurol Disord. 2004 Jun;3(3):169-79. doi: 10.2174/1568007043337409. Curr Drug Targets CNS Neurol Disord. 2004. PMID: 15180478 Review.
-
Distribution of extrasynaptic NMDA receptors on neurons.ScientificWorldJournal. 2012;2012:267120. doi: 10.1100/2012/267120. Epub 2012 Apr 30. ScientificWorldJournal. 2012. PMID: 22654580 Free PMC article. Review.
Cited by
-
Do specific NMDA receptor subunits act as gateways for addictive behaviors?Genes Brain Behav. 2017 Jan;16(1):118-138. doi: 10.1111/gbb.12348. Epub 2016 Nov 18. Genes Brain Behav. 2017. PMID: 27706932 Free PMC article. Review.
-
GluN2A versus GluN2B: twins, but quite different.Neurosci Bull. 2013 Dec;29(6):761-72. doi: 10.1007/s12264-013-1336-9. Epub 2013 Apr 20. Neurosci Bull. 2013. PMID: 23604599 Free PMC article. Review.
-
Chronic ethanol ingestion modulates proanxiety factors expressed in rat central amygdala.Alcohol. 2005 Jun;36(2):83-90. doi: 10.1016/j.alcohol.2005.07.004. Alcohol. 2005. PMID: 16396741 Free PMC article.
-
Glutamate plasticity woven through the progression to alcohol use disorder: a multi-circuit perspective.F1000Res. 2017 Mar 21;6:298. doi: 10.12688/f1000research.9609.1. eCollection 2017. F1000Res. 2017. PMID: 28413623 Free PMC article. Review.
-
Effects of topiramate and other anti-glutamatergic drugs on the acute intoxicating actions of ethanol in mice: modulation by genetic strain and stress.Neuropsychopharmacology. 2009 May;34(6):1454-66. doi: 10.1038/npp.2008.182. Epub 2008 Oct 8. Neuropsychopharmacology. 2009. PMID: 18843265 Free PMC article.
References
-
- Barria A, Malinow R (2002) Subunit-specific NMDA receptor trafficking to synapses. Neuron 35: 345-353. - PubMed
-
- Blanpied TA, Scott DB, Ehlers MD (2002) Dynamics and regulation of clathrin coats at specialized endocytic zones of dendrites and spines. Neuron 24: 435-449. - PubMed
-
- Carroll RC, Zukin RS (2002) NMDA-receptor trafficking and targeting: implications for synaptic transmission and plasticity. Trends Neurosci 25: 571-577. - PubMed
-
- Cebere A, Cebers G, Liljequist S (1999) Enhancement of NMDA-induced functional responses without concomitant NMDA receptor changes following chronic ethanol exposure in cerebellar granule cells. Naunyn Schmiedebergs Arch Pharmacol 360: 623-632. - PubMed
-
- Chandler LJ (2003) Ethanol and brain plasticity: receptors and molecular networks of the postsynaptic density as targets of ethanol. Pharmacol Ther 99: 311-326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- RR15776/RR/NCRR NIH HHS/United States
- AA09986/AA/NIAAA NIH HHS/United States
- F31 AA013355/AA/NIAAA NIH HHS/United States
- P50 AA010761/AA/NIAAA NIH HHS/United States
- AA00238/AA/NIAAA NIH HHS/United States
- K02 AA000238/AA/NIAAA NIH HHS/United States
- R01 AA009986/AA/NIAAA NIH HHS/United States
- R37 AA009986/AA/NIAAA NIH HHS/United States
- AA010761/AA/NIAAA NIH HHS/United States
- AA13355/AA/NIAAA NIH HHS/United States
- R01 AA010983/AA/NIAAA NIH HHS/United States
- AA010983/AA/NIAAA NIH HHS/United States
- R29 AA010983/AA/NIAAA NIH HHS/United States
LinkOut - more resources
Full Text Sources