

The upregulated expression of sonic hedgehog in motor neurons after rat facial nerve axotomy
- PMID: 15356205
- PMCID: PMC6729937
- DOI: 10.1523/JNEUROSCI.1784-04.2004
The upregulated expression of sonic hedgehog in motor neurons after rat facial nerve axotomy
Abstract
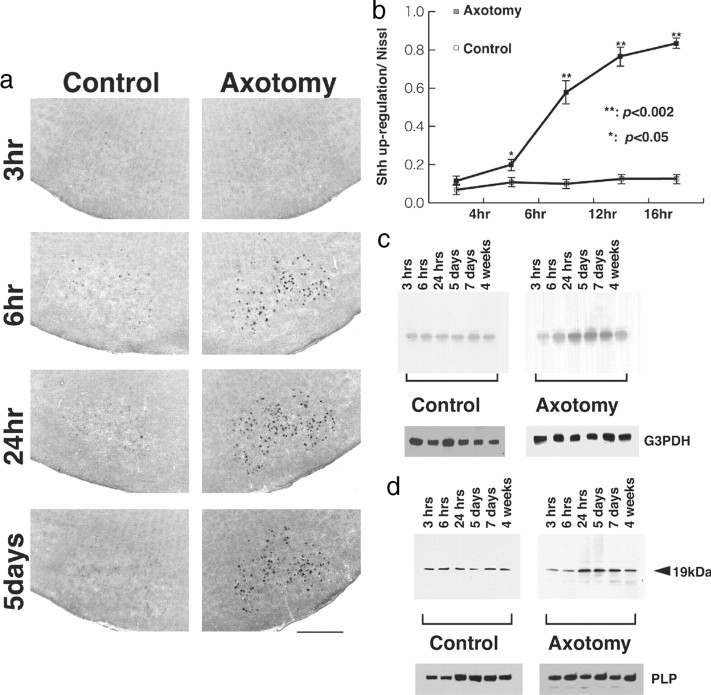
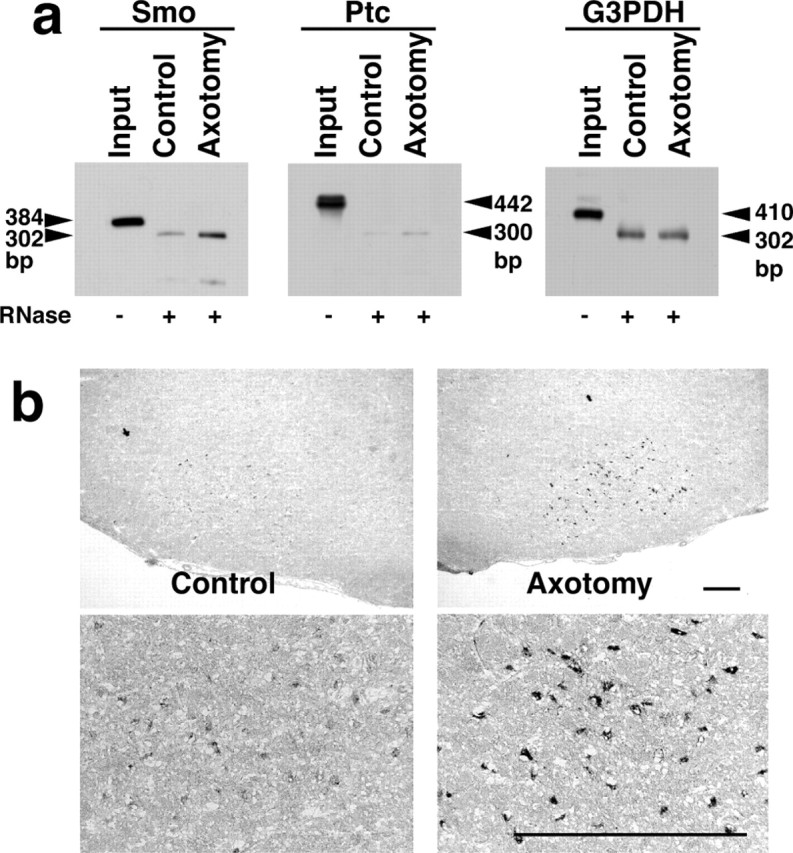
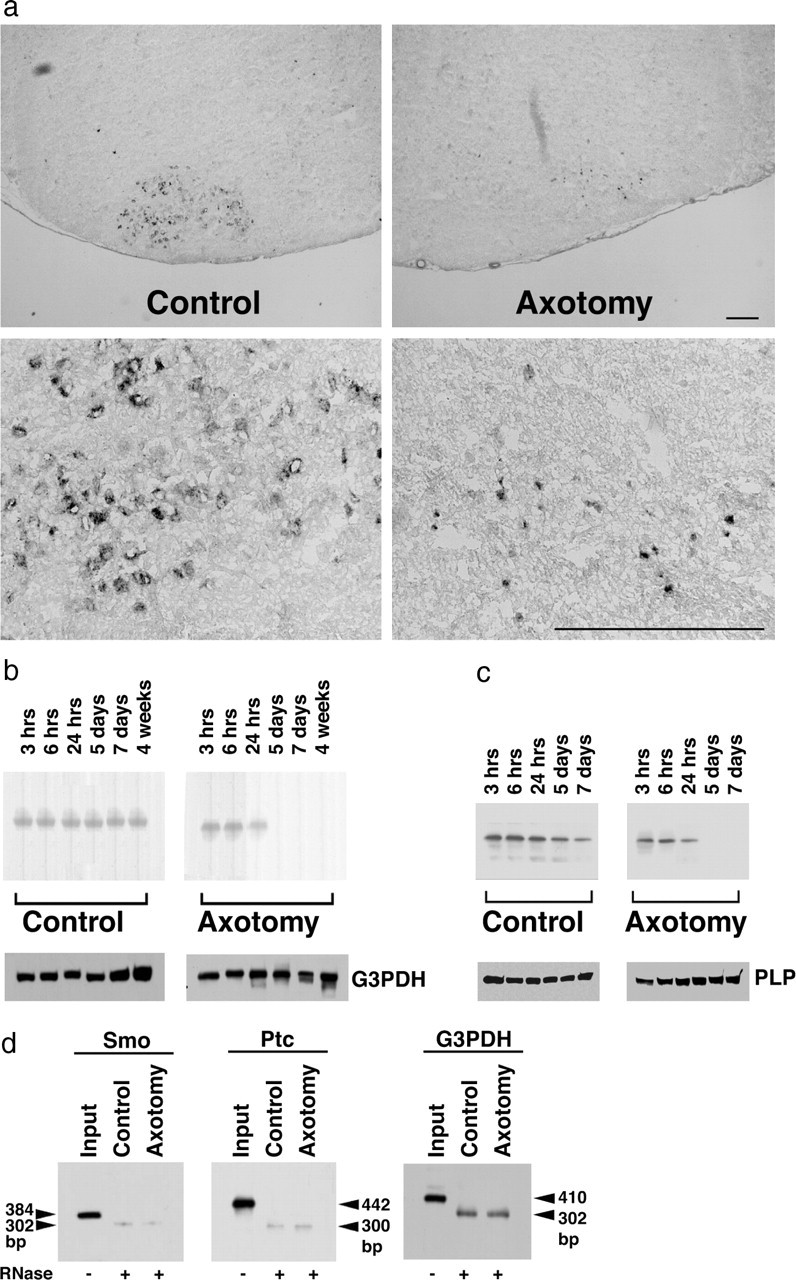
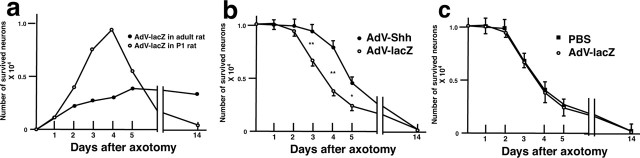
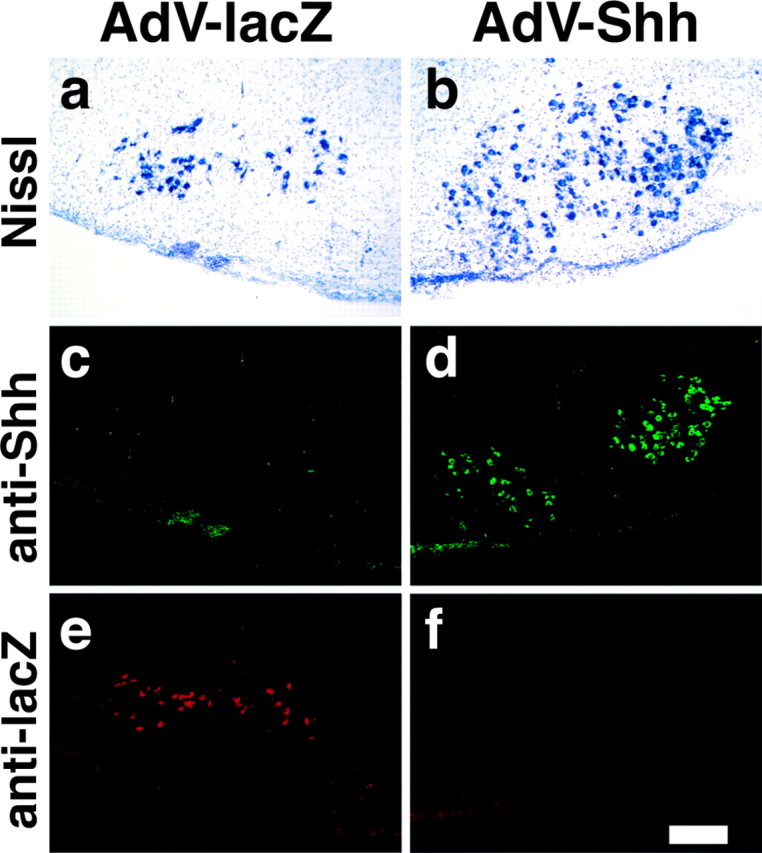
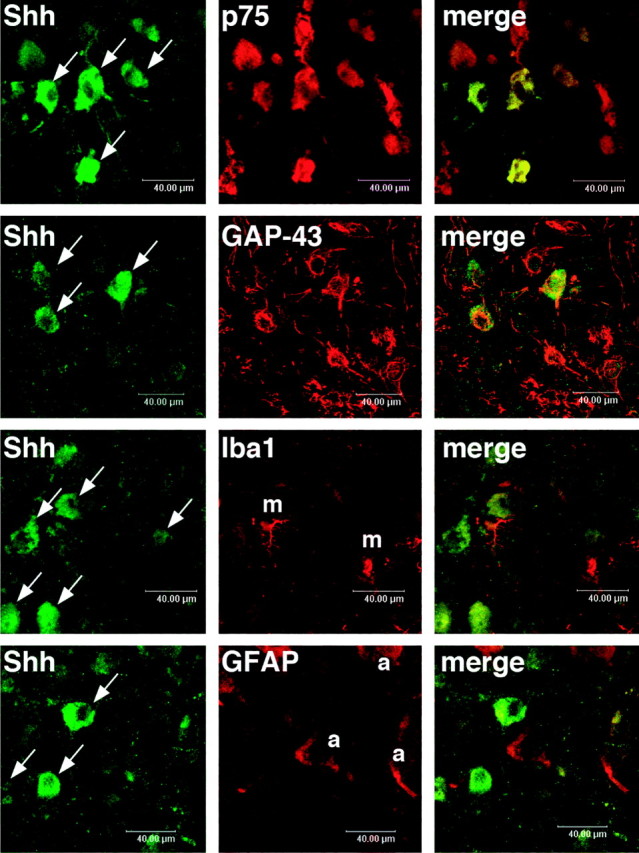
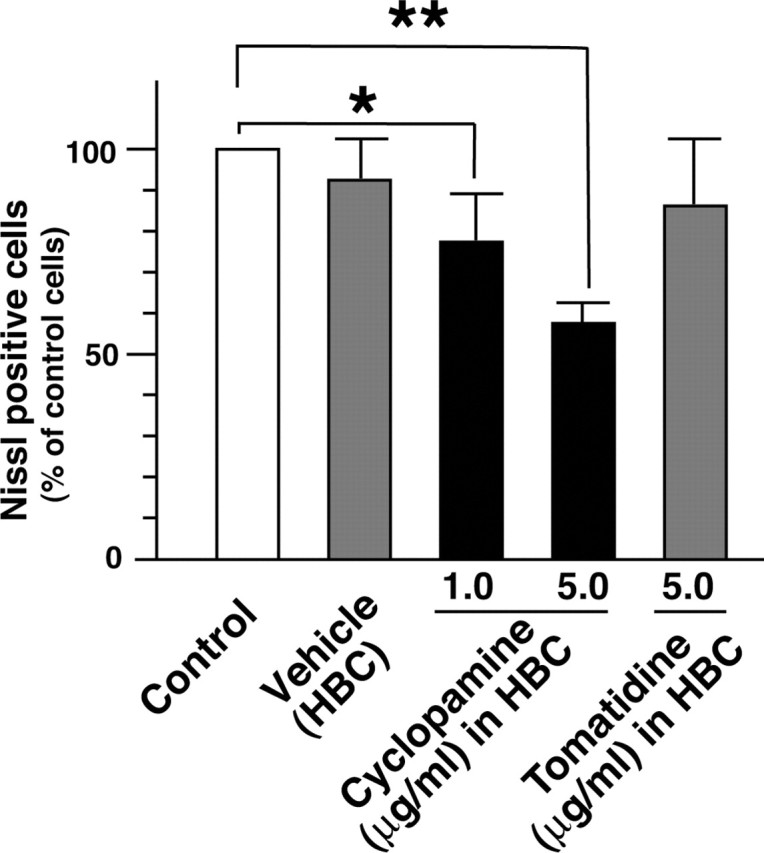
Nerve injury leads to the induction of a large number of genes to repair the damage and to restore synaptic transmission. We have attempted to identify molecules whose mRNA expression is altered in response to facial nerve axotomy. Here we report that facial nerve axotomy upregulates Sonic hedgehog (Shh) and its receptor Smoothened (Smo) in facial motor neurons of adult rats, whereas facial nerve axotomy does not upregulate mRNA of Shh or Smo in neonatal rats. We tested whether overexpression of Shh in facial motor neurons of axotomized neonatal rats may promote neuronal survival. Adenovirus-mediated overexpression of Shh, but not that of beta-galactosidase, transiently rescues axotomy-induced neuronal cell death for 3-5 d after axotomy. Finally, the pharmacological inhibitor of Shh signaling, cyclopamine, induces motor neuron death in adult rats after axotomy. These results suggest that Shh plays a regulatory role in nerve injury.
Figures
References
-
- Akazawa C, Nakamura Y, Sango K, Horie H, Kohsaka S (2004) Distribution of the galectin-1 mRNA in the rat nervous system: its transient upregulation in rat facial motor neurons after facial nerve axotomy. Neuroscience 125: 171-178. - PubMed
-
- Baumgartner BJ, Shine HD (1998) Permanent rescue of lesioned neonatal motoneurons and enhanced axonal regeneration by adenovirus-mediated expression of glial cell-line-derived neurotrophic factor. J Neurosci Res 54: 766-777. - PubMed
-
- Berman DM, Karhadkar SS, Hallahan AR, Pritchard JI, Eberhart CG, Watkins DN, Chen JK, Cooper MK, Taipale J, Olson JM, Beachy PA (2002) Medulloblastoma growth inhibition by hedgehog pathway blockade. Science 297: 1559-1561. - PubMed
-
- Briscoe J, Sussel L, Serup P, Hartigan-O'Connor D, Jessell TM, Rubenstein JL, Ericson J (1999) Homeobox gene Nkx2.2 and specification of neuronal identity by graded Sonic hedgehog signaling. Nature 398: 622-627. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous