

Rb is required for progression through myogenic differentiation but not maintenance of terminal differentiation
- PMID: 15364961
- PMCID: PMC2172111
- DOI: 10.1083/jcb.200403004
Rb is required for progression through myogenic differentiation but not maintenance of terminal differentiation
Abstract
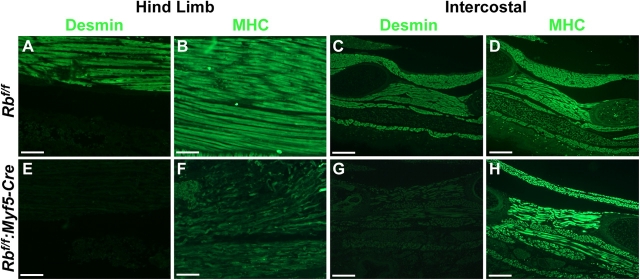
To investigate the requirement for pRb in myogenic differentiation, a floxed Rb allele was deleted either in proliferating myoblasts or after differentiation. Myf5-Cre mice, lacking pRb in myoblasts, died immediately at birth and exhibited high numbers of apoptotic nuclei and an almost complete absence of myofibers. In contrast, MCK-Cre mice, lacking pRb in differentiated fibers, were viable and exhibited a normal muscle phenotype and ability to regenerate. Induction of differentiation of Rb-deficient primary myoblasts resulted in high rates of apoptosis and a total inability to form multinucleated myotubes. Upon induction of differentiation, Rb-deficient myoblasts up-regulated myogenin, an immediate early marker of differentiation, but failed to down-regulate Pax7 and exhibited growth in low serum conditions. Primary myoblasts in which Rb was deleted after expression of differentiated MCK-Cre formed normal multinucleated myotubes that did not enter S-phase in response to serum stimulation. Therefore, Rb plays a crucial role in the switch from proliferation to differentiation rather than maintenance of the terminally differentiated state.
Figures









Similar articles
-
Regulation of Rb gene expression by an MBD2-interacting zinc finger protein MIZF during myogenic differentiation.Biochem Biophys Res Commun. 2004 Dec 17;325(3):653-9. doi: 10.1016/j.bbrc.2004.10.090. Biochem Biophys Res Commun. 2004. PMID: 15541338
-
Rescue of myogenic defects in Rb-deficient cells by inhibition of autophagy or by hypoxia-induced glycolytic shift.J Cell Biol. 2010 Oct 18;191(2):291-301. doi: 10.1083/jcb.201005067. Epub 2010 Oct 11. J Cell Biol. 2010. PMID: 20937698 Free PMC article.
-
Stable expression of antisense Rb-1 RNA inhibits terminal differentiation of mouse myoblast C2 cells.Exp Cell Res. 1998 Feb 25;239(1):40-9. doi: 10.1006/excr.1997.3880. Exp Cell Res. 1998. PMID: 9511723
-
CMF1-Rb interaction promotes myogenesis in avian skeletal myoblasts.Dev Dyn. 2008 May;237(5):1424-33. doi: 10.1002/dvdy.21544. Dev Dyn. 2008. PMID: 18425850 Free PMC article.
-
Restoring the Cell Cycle and Proliferation Competence in Terminally Differentiated Skeletal Muscle Myotubes.Cells. 2021 Oct 14;10(10):2753. doi: 10.3390/cells10102753. Cells. 2021. PMID: 34685732 Free PMC article. Review.
Cited by
-
A high-content, high-throughput siRNA screen identifies cyclin D2 as a potent regulator of muscle progenitor cell fusion and a target to enhance muscle regeneration.Hum Mol Genet. 2013 Aug 15;22(16):3283-95. doi: 10.1093/hmg/ddt184. Epub 2013 Apr 23. Hum Mol Genet. 2013. PMID: 23612904 Free PMC article.
-
Temporal activation of XRCC1-mediated DNA repair is essential for muscle differentiation.Cell Discov. 2016 Jan 12;2:15041. doi: 10.1038/celldisc.2015.41. eCollection 2016. Cell Discov. 2016. PMID: 27462438 Free PMC article.
-
Transcriptional control of stem cell fate by E2Fs and pocket proteins.Front Genet. 2015 Apr 28;6:161. doi: 10.3389/fgene.2015.00161. eCollection 2015. Front Genet. 2015. PMID: 25972892 Free PMC article. Review.
-
Retinoblastoma protein plays multiple essential roles in the terminal differentiation of Sertoli cells.Mol Endocrinol. 2009 Nov;23(11):1900-13. doi: 10.1210/me.2009-0184. Epub 2009 Oct 9. Mol Endocrinol. 2009. PMID: 19819985 Free PMC article.
-
Interplay between the retinoblastoma protein and LEK1 specifies stem cells toward the cardiac lineage.EMBO J. 2005 May 4;24(9):1750-61. doi: 10.1038/sj.emboj.7600652. Epub 2005 Apr 21. EMBO J. 2005. PMID: 15861132 Free PMC article.
References
-
- Bergstrom, D.A., B.H. Penn, A. Strand, R.L. Perry, M.A. Rudnicki, and S.J. Tapscott. 2002. Promoter-specific regulation of MyoD binding and signal transduction cooperate to pattern gene expression. Mol. Cell. 9:587–600. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
