

Nuclear export of the nonenveloped parvovirus virion is directed by an unordered protein signal exposed on the capsid surface
- PMID: 15367635
- PMCID: PMC516424
- DOI: 10.1128/JVI.78.19.10685-10694.2004
Nuclear export of the nonenveloped parvovirus virion is directed by an unordered protein signal exposed on the capsid surface
Abstract
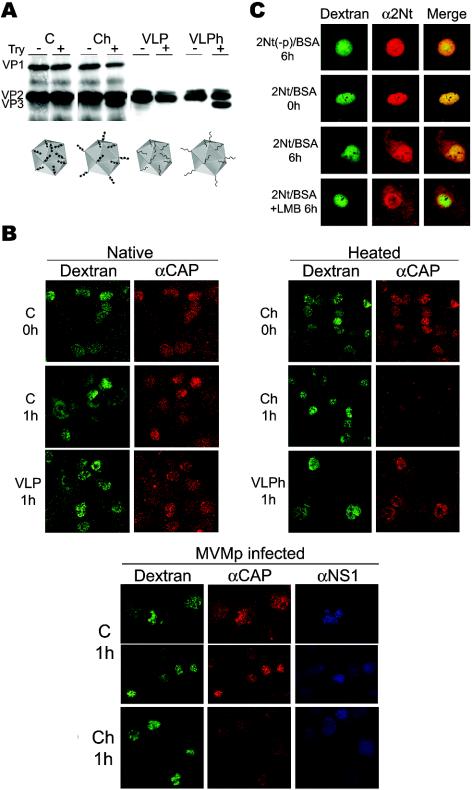
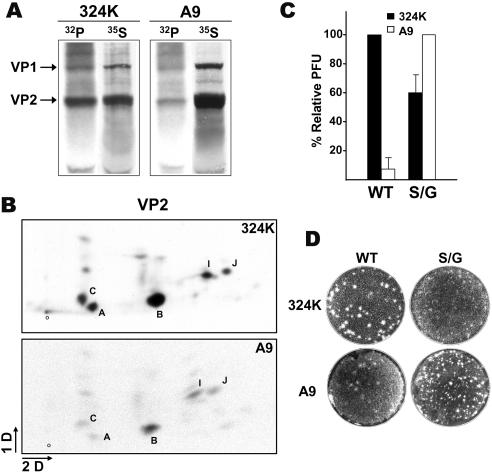
It is uncertain whether nonenveloped karyophilic virus particles may actively traffic from the nucleus outward. The unordered amino-terminal domain of the VP2 major structural protein (2Nt) of the icosahedral parvovirus minute virus of mice (MVM) is internal in empty capsids, but it is exposed outside of the shell through the fivefold axis of symmetry in virions with an encapsidated single-stranded DNA genome, as well as in empty capsids subjected to a heat-induced structural transition. In productive infections of transformed and normal fibroblasts, mature MVM virions were found to efficiently exit from the nucleus prior to cell lysis, in contrast to the extended nuclear accumulation of empty capsids. Newly formed mutant viruses lacking the three phosphorylated serine residues of 2Nt were hampered in their exit from the human transformed NB324K nucleus, in correspondence with the capacity of 2Nt to drive microinjected phosphorylated heated capsids out of the nucleus. However, in normal mouse A9 fibroblasts, in which the MVM capsid was phosphorylated at similar sites but with a much lower rate, the nuclear exit of virions and microinjected capsids harboring exposed 2Nt required the infection process and was highly sensitive to inhibition of the exportin CRM1 in the absence of a demonstrable interaction. Thus, the MVM virion exits the nucleus by accessing nonconventional export pathways relying on cell physiology that can be intensified by infection but in which the exposure of 2Nt remains essential for transport. The flexible 2Nt nuclear transport signal may illustrate a common structural solution used by nonenveloped spherical viruses to propagate in undamaged host tissues.
Figures





References
-
- Agbandje-McKenna, M., A. LLamas-Saiz, F. Wang, P. Tattersall, and M. G. Rossmann. 1998. Functional implications of the structure of the murine parvovirus minute virus of mice. Structure 6:1369-1381. - PubMed
-
- Boyle, W. J., P. van der Geer, and T. Hunter. 1991. Phosphopeptide mapping and phosphoamino acid analysis by two-dimensional separation on thin-layer cellulose plates. Methods Enzymol. 201:110-149. - PubMed
-
- Brown, V. M., E. Y. Krynetski, N. F. Krynetskaia, D. Grieger, S. T. Mukatira, K. G. Murti, C. A. Slaughter, H.-W. Park, and W. E. Evans. 2004. A novel crm1-mediated nuclear export signal governs nuclear accumulation of glyceraldehyde-3-phosphate dehydrogenase following genotoxic stress. J. Biol. Chem. 279:5984-5992. - PubMed
-
- Brownstein, D. G., A. L. Smith, R. O. Jacoby, E. A. Johnson, G. Hansen, and P. Tattersall. 1991. Pathogenesis of infection with a virulent allotropic variant of minute virus of mice and regulation by host genotype. Lab. Investig. 65:357-363. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
