

The ubiquitin-conjugating enzyme UBCH7 acts as a coactivator for steroid hormone receptors
- PMID: 15367689
- PMCID: PMC516762
- DOI: 10.1128/MCB.24.19.8716-8726.2004
The ubiquitin-conjugating enzyme UBCH7 acts as a coactivator for steroid hormone receptors
Abstract
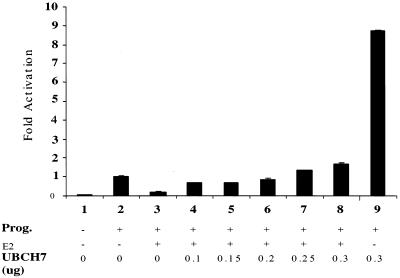
We investigated the role of the ubiquitin-conjugating enzyme UBCH7 in nuclear receptor transactivation. Using transient transfection assays, we demonstrated that UBCH7 modulates the transcriptional activity of progesterone receptor (PR) and glucocorticoid, androgen, and retinoic acid receptors in a hormone-dependent manner and that the ubiquitin conjugation activity of UBCH7 is required for its ability to potentiate transactivation by steroid hormone receptors (SHR). However, UBCH7 showed no significant effect on the transactivation functions of p53 and VP-16 activation domain. Depletion of endogenous UBCH7 protein by small interfering RNAs suggests that UBCH7 is required for the proper function of SHR. Furthermore, a chromatin immunoprecipitation assay demonstrated the hormone-dependent recruitment of UBCH7 onto estrogen receptor- and PR-responsive promoters. Additionally, we show that UBCH7 and E6-associated protein (E6-AP) synergistically enhance PR transactivation. We also demonstrate that UBCH7 interacts with steroid receptor coactivator 1 (SRC-1) and that UBCH7 coactivation function is dependent on SRC-1. Taken together, our results reveal the possible role of UBCH7 in steroid receptor transactivation and provide insights into the mechanism of action of UBCH7 in receptor function.
Figures











Similar articles
-
WW domain binding protein-2, an E6-associated protein interacting protein, acts as a coactivator of estrogen and progesterone receptors.Mol Endocrinol. 2006 Oct;20(10):2343-54. doi: 10.1210/me.2005-0533. Epub 2006 Jun 13. Mol Endocrinol. 2006. PMID: 16772533
-
Ligand-controlled interaction of histone acetyltransferase binding to ORC-1 (HBO1) with the N-terminal transactivating domain of progesterone receptor induces steroid receptor coactivator 1-dependent coactivation of transcription.Mol Endocrinol. 2006 Sep;20(9):2122-40. doi: 10.1210/me.2005-0149. Epub 2006 Apr 27. Mol Endocrinol. 2006. PMID: 16645042
-
E1A-mediated repression of progesterone receptor-dependent transactivation involves inhibition of the assembly of a multisubunit coactivation complex.Mol Cell Biol. 2000 Mar;20(6):2138-46. doi: 10.1128/MCB.20.6.2138-2146.2000. Mol Cell Biol. 2000. PMID: 10688660 Free PMC article.
-
Role of co-activators and co-repressors in the mechanism of steroid/thyroid receptor action.Recent Prog Horm Res. 1997;52:141-64; discussion 164-5. Recent Prog Horm Res. 1997. PMID: 9238851 Review.
-
Cdc25B as a steroid receptor coactivator.Vitam Horm. 2004;68:231-56. doi: 10.1016/S0083-6729(04)68008-3. Vitam Horm. 2004. PMID: 15193457 Review.
Cited by
-
Links between oestrogen receptor activation and proteolysis: relevance to hormone-regulated cancer therapy.Nat Rev Cancer. 2014 Jan;14(1):26-38. doi: 10.1038/nrc3622. Nat Rev Cancer. 2014. PMID: 24505618 Review.
-
Ligand-dependent degradation of SRC-1 is pivotal for progesterone receptor transcriptional activity.Mol Endocrinol. 2011 Mar;25(3):394-408. doi: 10.1210/me.2010-0458. Epub 2011 Jan 27. Mol Endocrinol. 2011. PMID: 21273440 Free PMC article.
-
Estrogen inhibits transforming growth factor beta signaling by promoting Smad2/3 degradation.J Biol Chem. 2010 May 7;285(19):14747-55. doi: 10.1074/jbc.M109.093039. Epub 2010 Mar 5. J Biol Chem. 2010. PMID: 20207742 Free PMC article.
-
Ubiquitin conjugating enzyme E2 L3 promoted tumor growth of NSCLC through accelerating p27kip1 ubiquitination and degradation.Oncotarget. 2017 Aug 24;8(48):84193-84203. doi: 10.18632/oncotarget.20449. eCollection 2017 Oct 13. Oncotarget. 2017. PMID: 29137415 Free PMC article.
-
Targeting the binding function 3 (BF3) site of the human androgen receptor through virtual screening.J Med Chem. 2011 Dec 22;54(24):8563-73. doi: 10.1021/jm201098n. Epub 2011 Nov 18. J Med Chem. 2011. PMID: 22047606 Free PMC article.
References
-
- Allan, G. F., X. Leng, S. Y. Tsai, N. L. Weigel, D. P. Edwards, M. J. Tsai, and B. W. O'Malley. 1992. Hormone and antihormone induce distinct conformational changes which are central to steroid receptor activation. J. Biol. Chem. 267:19513-19520. - PubMed
-
- Aranda, A., and A. Pascual. 2001. Nuclear hormone receptors and gene expression. Physiol. Rev. 81:1269-1304. - PubMed
-
- Carson-Jurica, M. A., W. T. Schrader, and B. W. O'Malley. 1990. Steroid receptor family: structure and functions. Endocr. Rev. 11:201-220. - PubMed
-
- Chen, D., H. Ma, H. Hong, S. S. Koh, S. M. Huang, B. T. Schurter, D. W. Aswad, and M. R. Stallcup. 1999. Regulation of transcription by a protein methyltransferase. Science 284:2174-2177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous