

Essential role of BCL9-2 in the switch between beta-catenin's adhesive and transcriptional functions
- PMID: 15371335
- PMCID: PMC517514
- DOI: 10.1101/gad.317604
Essential role of BCL9-2 in the switch between beta-catenin's adhesive and transcriptional functions
Abstract
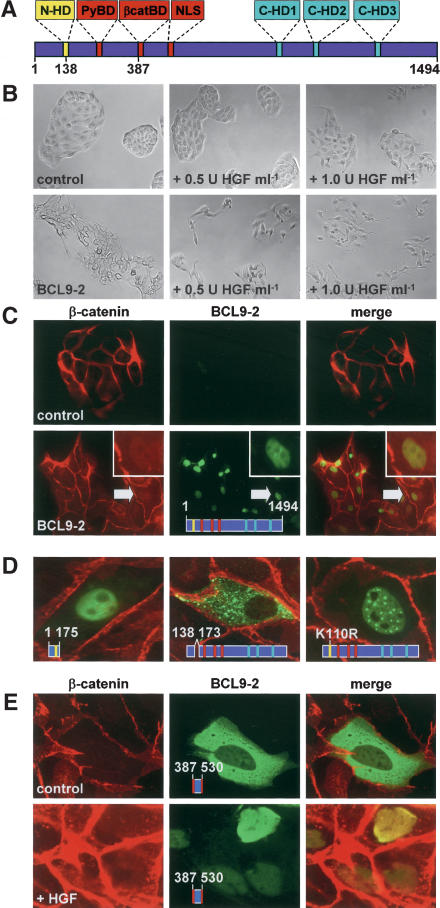
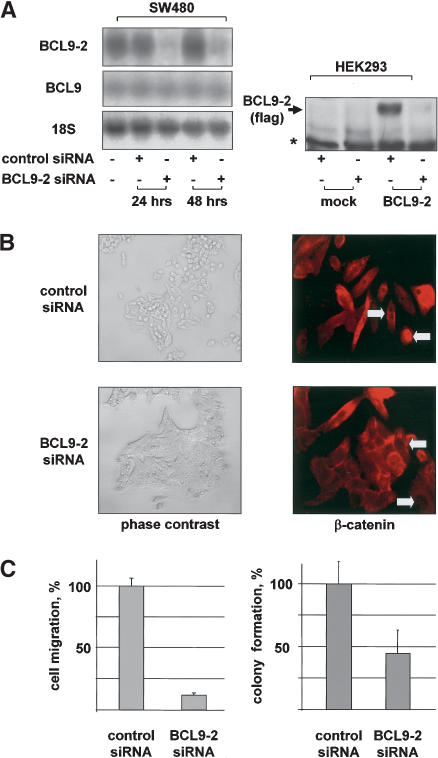
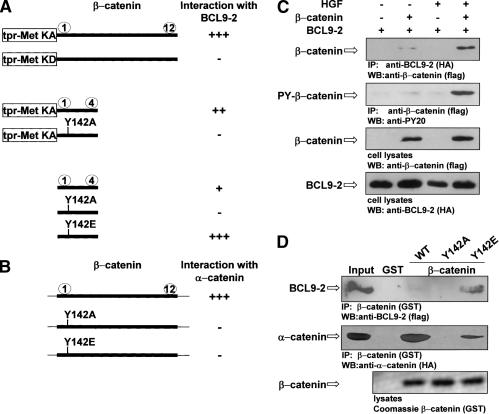
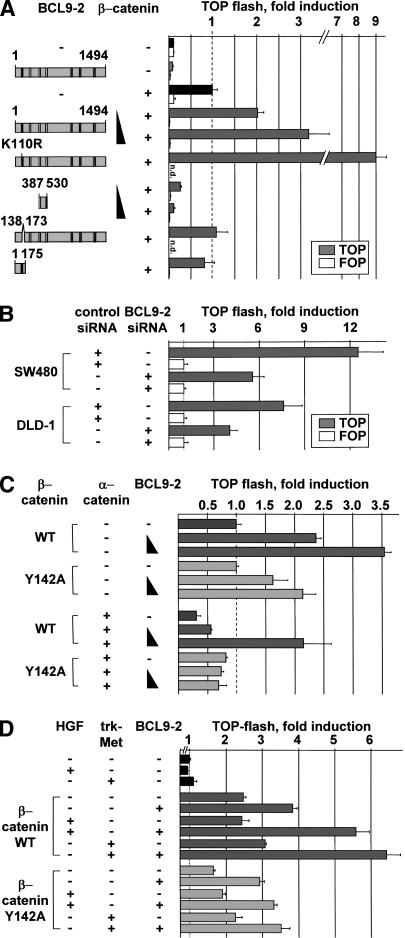
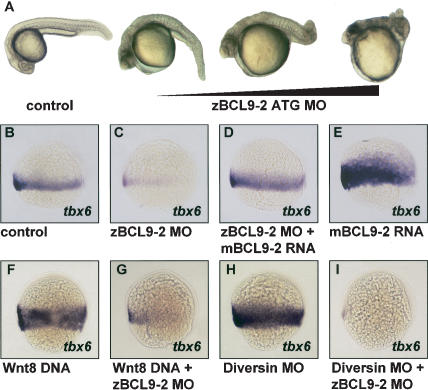
beta-Catenin controls both cadherin-mediated cell adhesion and activation of Wnt target genes. We demonstrate here that the beta-catenin-binding protein BCL9-2, a homolog of the human proto-oncogene product BCL9, induces epithelial-mesenchymal transitions of nontransformed cells and increases beta-catenin-dependent transcription. RNA interference of BCL9-2 in carcinoma cells induces an epithelial phenotype and translocates beta-catenin from the nucleus to the cell membrane. The switch between beta-catenin's adhesive and transcriptional functions is modulated by phosphorylation of Tyr 142 of beta-catenin, which favors BCL9-2 binding and precludes interaction with alpha-catenin. During zebrafish embryogenesis, BCL9-2 acts in the Wnt8-signaling pathway and regulates mesoderm patterning.
Figures
References
-
- Aberle H., Schwartz, H., Hoschuetzky, H., and Kemler, R. 1996. Single amino acid substitutions in proteins of the armadillo gene family abolish their binding to α-catenin. J. Biol. Chem. 271: 1520-1526. - PubMed
-
- Behrens J., Vakaet, L., Friis, R., Winterhager, E., Van Roy, F., Mareel, M.M., and Birchmeier, W. 1993. Loss of epithelial differentiation and gain of invasiveness correlates with tyrosine phosphorylation of the E-cadherin/β-catenin complex in cells transformed with a temperature-sensitive v-SRC gene. J. Cell Biol. 120: 757-766. - PMC - PubMed
-
- Behrens J., von Kries, J.P., Kuhl, M., Bruhn, L., Wedlich, D., Grosschedl, R., and Birchmeier, W. 1996. Functional interaction of β-catenin with the transcription factor LEF-1. Nature 382: 638-642. - PubMed
-
- Behrens J., Jerchow, B.A., Wurtele, M., Grimm, J., Asbrand, C., Wirtz, R., Kuhl, M., Wedlich, D., and Birchmeier, W. 1998. Functional interaction of an axin homolog, conductin, with β-catenin, APC, and GSK3β. Science 280: 596-599. - PubMed
-
- Bienz M. and Clevers, H. 2000. Linking colorectal cancer to Wnt signaling. Cell 103: 311-320. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous