

Reversible histone acetylation and deacetylation mediate genome-wide, promoter-dependent and locus-specific changes in gene expression during plant development
- PMID: 15371352
- PMCID: PMC1448893
- DOI: 10.1534/genetics.104.033142
Reversible histone acetylation and deacetylation mediate genome-wide, promoter-dependent and locus-specific changes in gene expression during plant development
Abstract
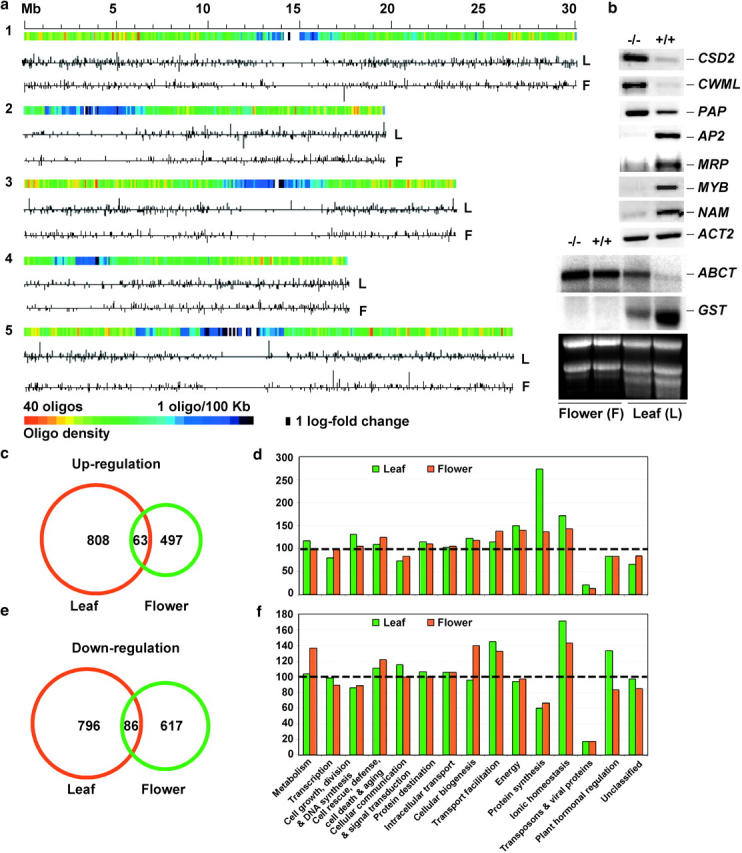
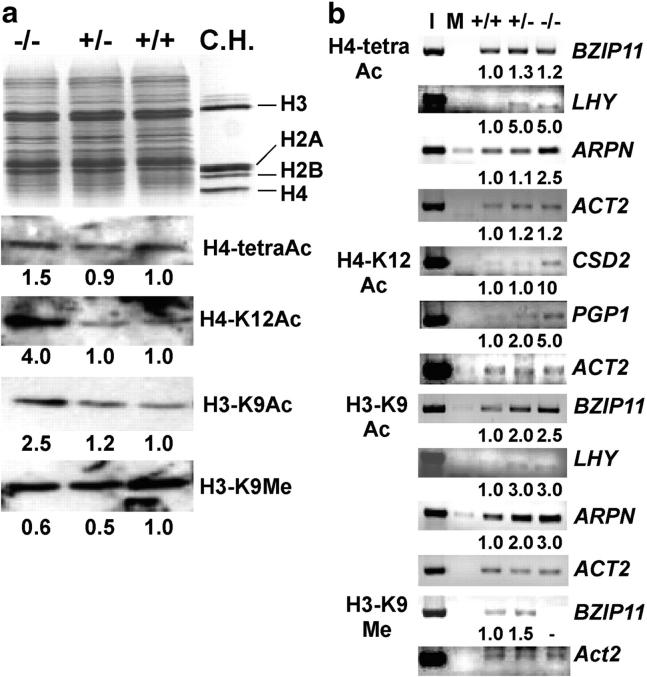
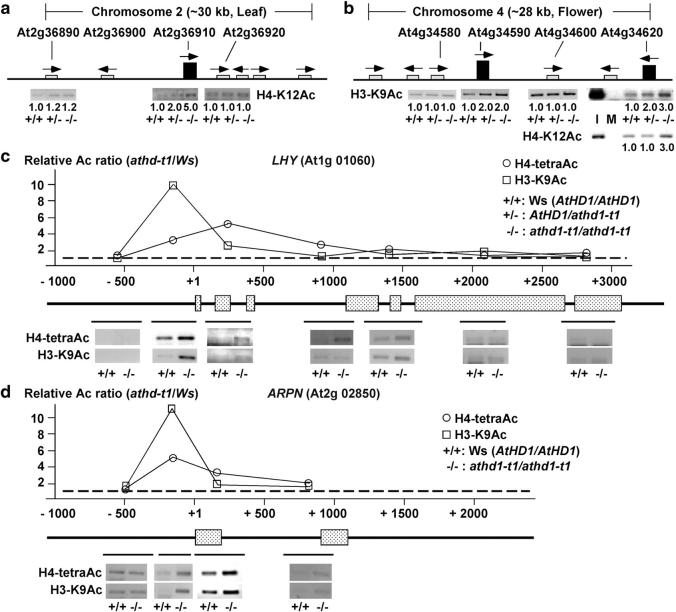
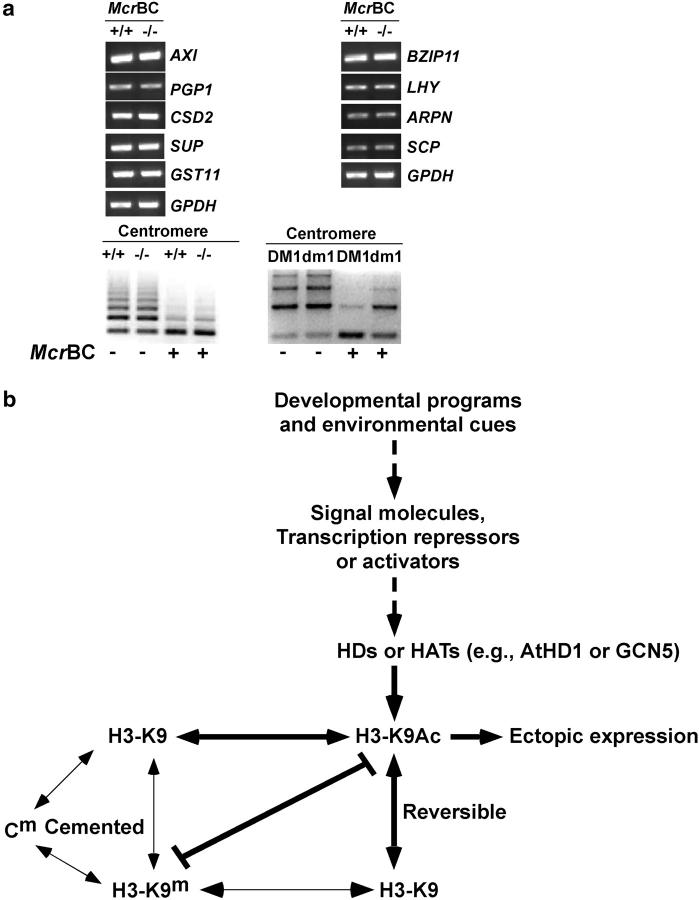
Histone acetylation and deacetylation activate or repress transcription, yet the physiological relevance of reversible changes in chromatin structure and gene expression is poorly understood. We have shown that disrupting the expression of AtHD1 that encodes a putative Arabidopsis thaliana histone deacetylase induces a variety of developmental abnormalities. However, causal effects of the AtHD1 disruption on chromatin structure and gene expression are unknown. Using Arabidopsis spotted oligo-gene microarray analysis, here we report that >7% of the transcriptome was up- or downregulated in A. thaliana plants containing a T-DNA insertion in AtHD1 (athd1-t1), indicating that AtHD1 provides positive and negative control of transcriptional regulation. Remarkably, genes involved in ionic homeostasis and protein synthesis were ectopically expressed, whereas genes in ionic homeostasis, protein transport, and plant hormonal regulation were repressed in athd1-t1 leaves or flowers, suggesting a role of AtHD1 in developmental and environmental regulation of gene expression. Moreover, defective AtHD1 induced site-specific and reversible acetylation changes in H3-Lys9, H4-Lys12, and H4 tetra-lysines (residues 5, 8, 12, and 16) in homozygous recessive and heterozygous plants. Transcriptional activation was locus specific and often associated with specific acetylation sites in the vicinity of promoters, whereas gene repression did not correlate with changes in histone acetylation or correlated directly with H3-Lys9 methylation but not with DNA methylation. The data suggest that histone acetylation and deacetylation are promoter dependent, locus specific, and genetically reversible, which provides a general mechanism for reversible gene regulation responsive to developmental and environmental changes.
Figures
References
-
- Alland, L., R. Muhle, H. Hou, Jr., J. Potes, L. Chin et al., 1997. Role for N-CoR and histone deacetylase in Sin3-mediated transcriptional repression. Nature 387: 49–55. - PubMed
-
- Arabidopsis Genome Initiative, 2000. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408: 796–815. - PubMed
-
- Ausin, I., C. Alonso-Blanco, J. A. Jarillo, L. Ruiz-Garcia and J. M. Martinez-Zapater, 2004. Regulation of flowering time by FVE, a retinoblastoma-associated protein. Nat. Genet. 36: 162–166. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
