

Nonlinear response properties of combination-sensitive electrosensory neurons in the midbrain of Gymnarchus niloticus
- PMID: 15371504
- PMCID: PMC6729786
- DOI: 10.1523/JNEUROSCI.2021-04.2004
Nonlinear response properties of combination-sensitive electrosensory neurons in the midbrain of Gymnarchus niloticus
Abstract
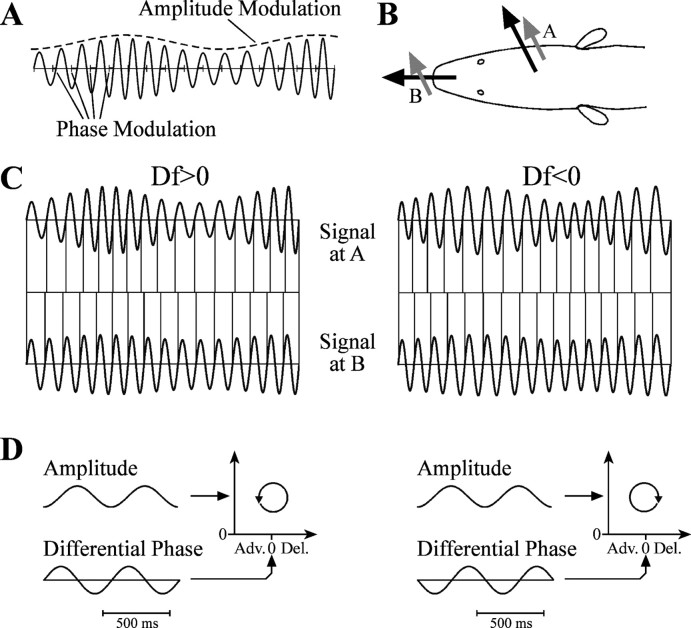
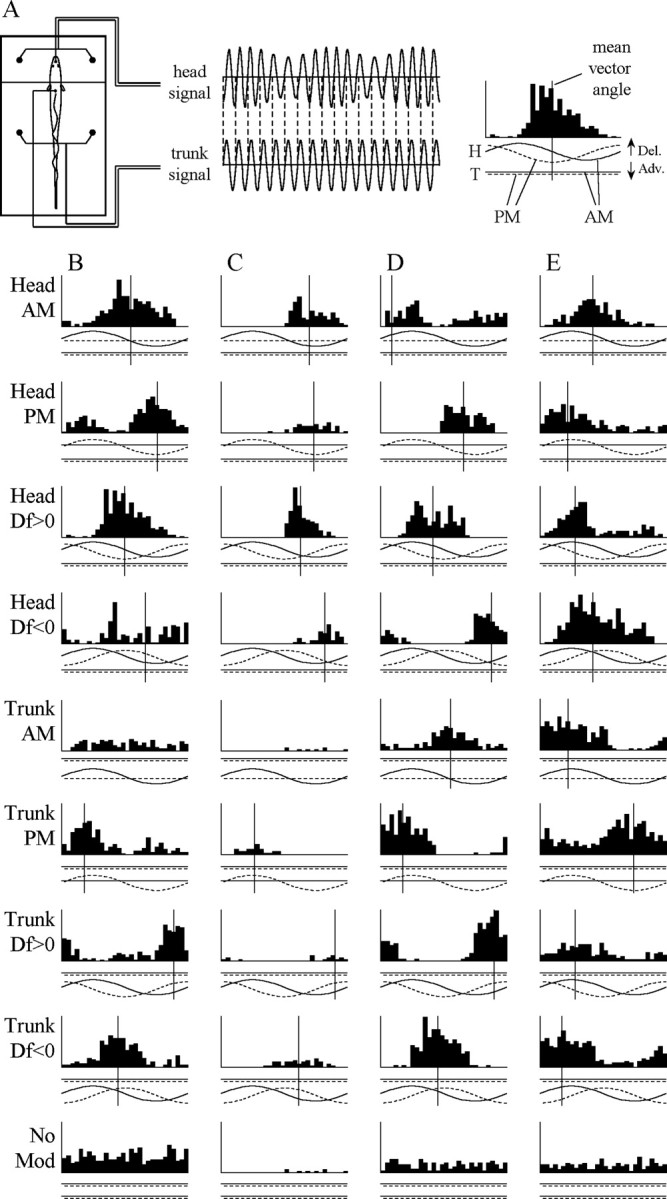
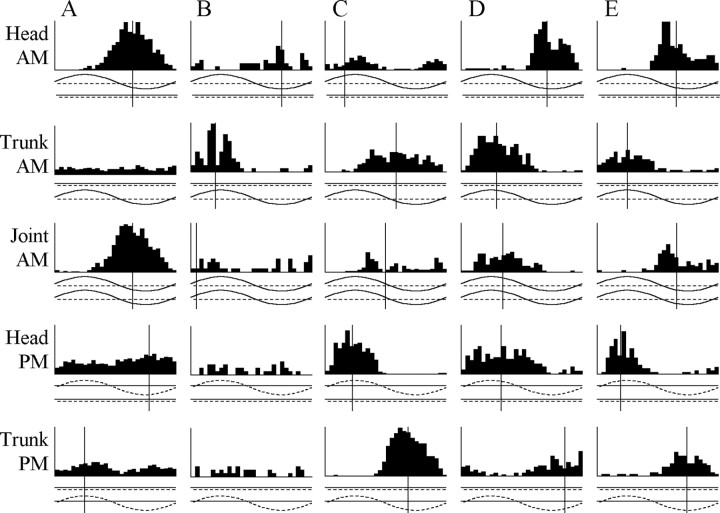
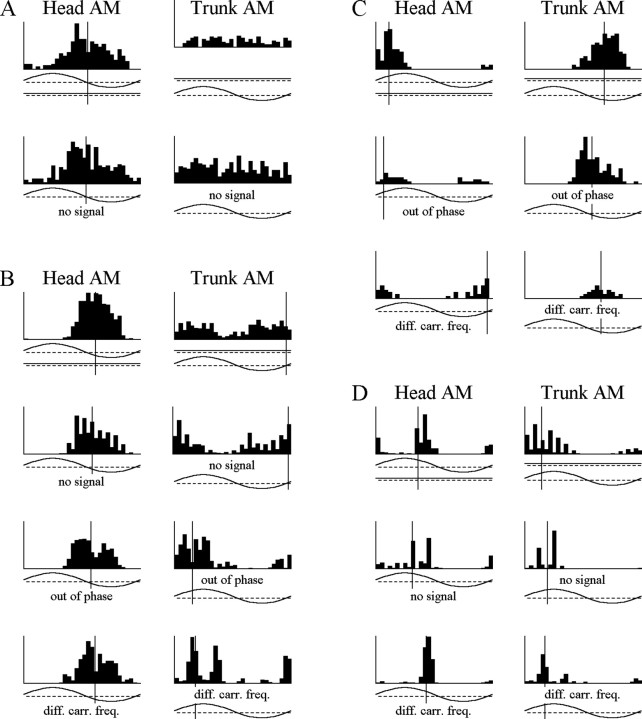
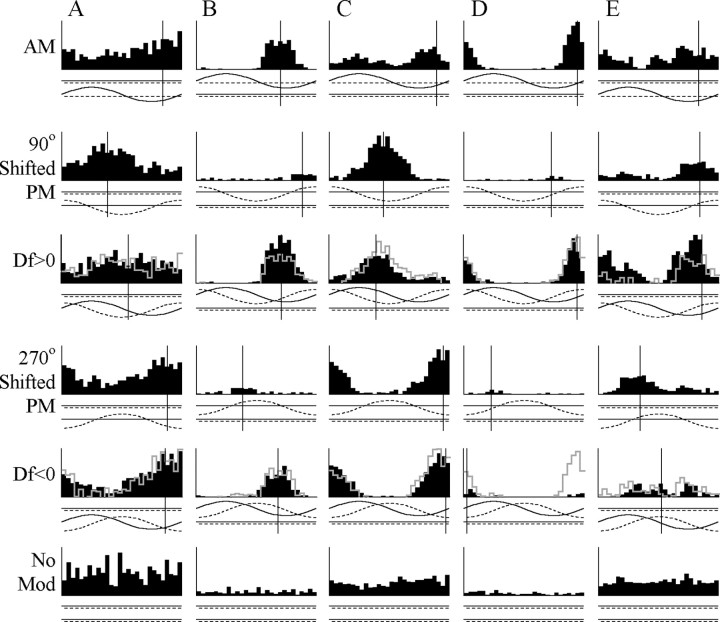
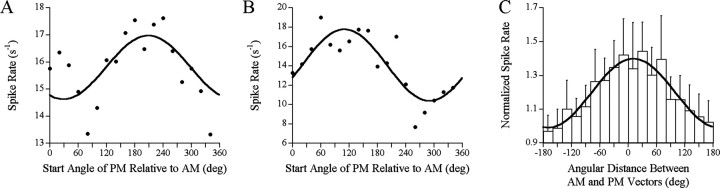
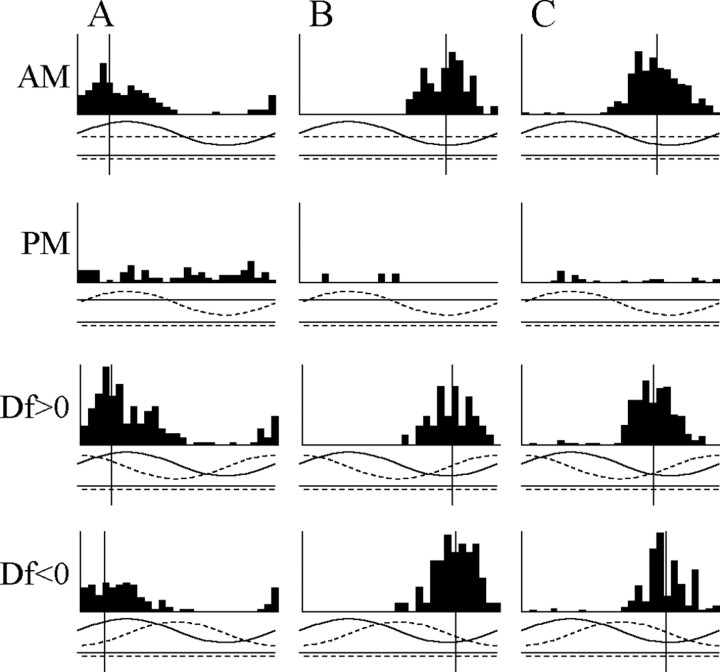
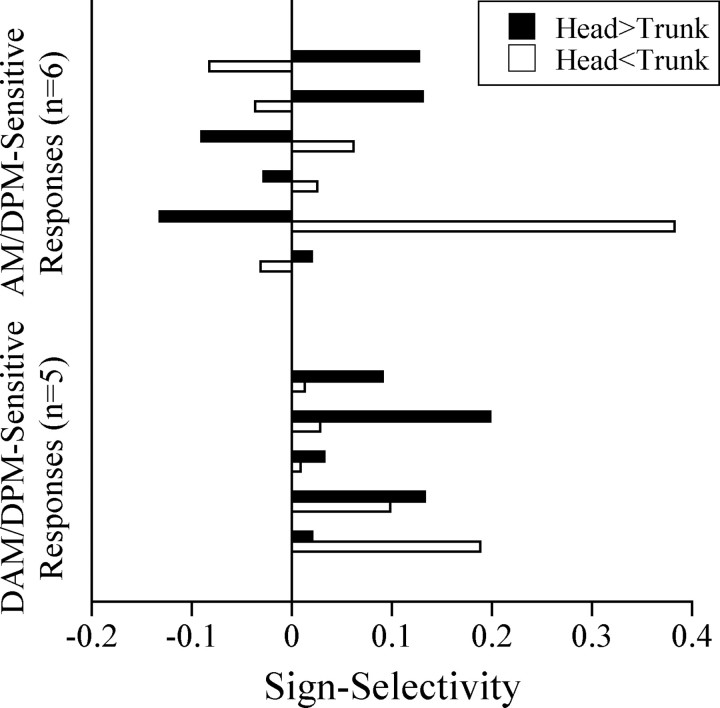
The jamming avoidance response of the weakly electric fish Gymnarchus niloticus relies on determining the sign of the frequency difference (Df) between the fish's own electric organ discharge (EOD) and that of a neighbor, which is achieved by comparing modulations in amplitude (AM) and phase (PM) that result from the summation of their EODs. These two stimulus features are processed in separate pathways that converge in the torus semicircularis on combination-sensitive neurons, many of which are selective for the sign of Df. We recorded extracellular single-unit responses to independent stimulation with AM and PM and combined AM-PM stimulation to determine how sign selectivity is established. Responses to AM and PM frequently summated nonlinearly, leading to sign-selective responses as a result of facilitation to the preferred sign of Df and/or suppression to the nonpreferred sign of Df. Facilitation typically occurred when responses to AM and PM were aligned, whereas suppression typically occurred when they were offset. By experimentally manipulating the degree of alignment between these two responses, we found that the summed response was dependent on their relative timing. In addition, we found a unique class of units that were sensitive to differences in amplitude between two body surfaces. This sensitivity rendered such units immune to the problem of orientation ambiguity, in which the sign selectivity of a single neuron reverses with changes in stimulus orientation. We discuss potential synaptic mechanisms for driving nonlinear responses in these and other combination-sensitive neurons.
Figures
Similar articles
-
Stimulus selectivity is enhanced by voltage-dependent conductances in combination-sensitive neurons.J Neurophysiol. 2006 Dec;96(6):3362-77. doi: 10.1152/jn.00839.2006. Epub 2006 Sep 27. J Neurophysiol. 2006. PMID: 17005607
-
Emergence of temporal-pattern sensitive neurons in the midbrain of weakly electric fish Gymnarchus niloticus.J Physiol Paris. 2002 Sep-Dec;96(5-6):531-7. doi: 10.1016/S0928-4257(03)00008-1. J Physiol Paris. 2002. PMID: 14692500
-
Stimulus discrimination in the diencephalon of Eigenmannia: the emergence and sharpening of a sensory filter.J Comp Physiol A. 1988 Apr;162(6):747-57. doi: 10.1007/BF00610964. J Comp Physiol A. 1988. PMID: 3397918
-
Perception and coding of envelopes in weakly electric fishes.J Exp Biol. 2013 Jul 1;216(Pt 13):2393-402. doi: 10.1242/jeb.082321. J Exp Biol. 2013. PMID: 23761464 Free PMC article. Review.
-
The neuroethology of electrocommunication: how signal background influences sensory encoding and behaviour in Apteronotus leptorhynchus.J Physiol Paris. 2013 Jan-Apr;107(1-2):13-25. doi: 10.1016/j.jphysparis.2012.07.001. Epub 2012 Sep 5. J Physiol Paris. 2013. PMID: 22981958 Review.
Cited by
-
Glycinergic "inhibition" mediates selective excitatory responses to combinations of sounds.J Neurosci. 2008 Jan 2;28(1):80-90. doi: 10.1523/JNEUROSCI.3572-07.2008. J Neurosci. 2008. PMID: 18171925 Free PMC article.
-
Temporal features of spectral integration in the inferior colliculus: effects of stimulus duration and rise time.J Neurophysiol. 2009 Jul;102(1):167-80. doi: 10.1152/jn.91300.2008. Epub 2009 Apr 29. J Neurophysiol. 2009. PMID: 19403742 Free PMC article.
-
From stimulus estimation to combination sensitivity: encoding and processing of amplitude and timing information in parallel, convergent sensory pathways.J Comput Neurosci. 2008 Aug;25(1):1-24. doi: 10.1007/s10827-007-0062-6. Epub 2008 Jan 5. J Comput Neurosci. 2008. PMID: 18176850 Free PMC article.
-
Ambiguous encoding of stimuli by primary sensory afferents causes a lack of independence in the perception of multiple stimulus attributes.J Neurosci. 2006 Sep 6;26(36):9173-83. doi: 10.1523/JNEUROSCI.1513-06.2006. J Neurosci. 2006. PMID: 16957074 Free PMC article.
-
Encoding and processing biologically relevant temporal information in electrosensory systems.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2006 Jun;192(6):625-35. doi: 10.1007/s00359-006-0102-0. Epub 2006 Feb 1. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2006. PMID: 16450118 Review.
References
-
- Bass AH, Hopkins CD (1982) Comparative aspects of brain organization of an African wave electric fish, Gymnarchus niloticus J Morphol 174: 313-334. - PubMed
-
- Batschelet E (1981) Circular statistics in biology, pp 54-58. New York: Academic.
-
- Bullock TH, Behrend K, Heiligenberg W (1975) Comparison of the jamming avoidance responses in gymnotoid and gymnarchid electric fish: a case of convergent evolution of behavior and its sensory basis. J Comp Physiol [A] 103: 97-121.
-
- Bullock TH, Bodznick DA, Northcutt RG (1983) The phylogenetic distribution of electroreception: evidence for convergent evolution of a primitive vertebrate sense modality. Brain Res Brain Res Rev 6: 25-46. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous