

PchC thioesterase optimizes nonribosomal biosynthesis of the peptide siderophore pyochelin in Pseudomonas aeruginosa
- PMID: 15375116
- PMCID: PMC516611
- DOI: 10.1128/JB.186.19.6367-6373.2004
PchC thioesterase optimizes nonribosomal biosynthesis of the peptide siderophore pyochelin in Pseudomonas aeruginosa
Abstract
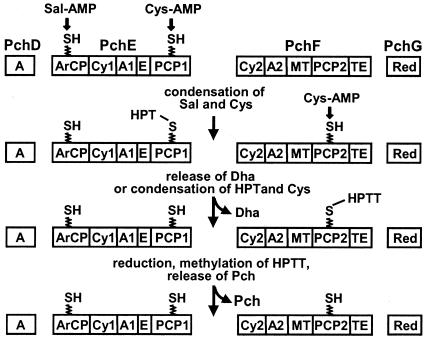
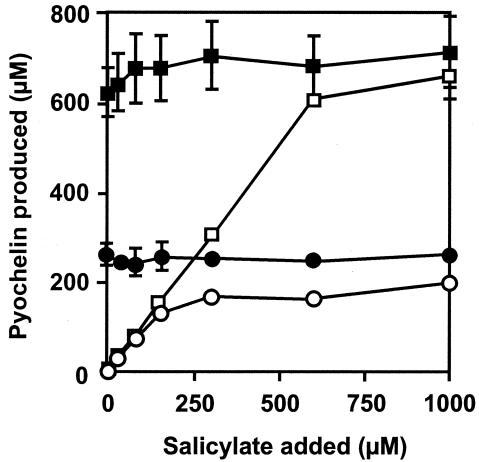
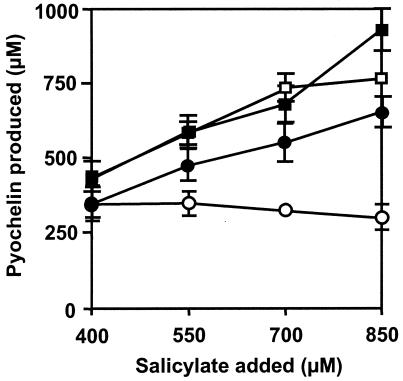
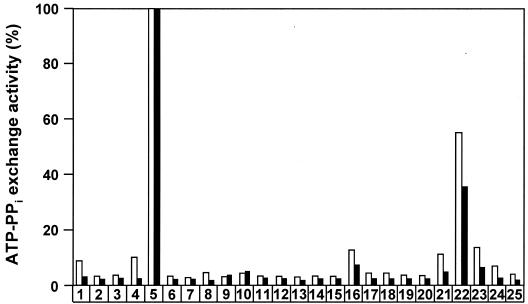
In Pseudomonas aeruginosa, the antibiotic dihydroaeruginoate (Dha) and the siderophore pyochelin are produced from salicylate and cysteine by a thiotemplate mechanism involving the peptide synthetases PchE and PchF. A thioesterase encoded by the pchC gene was found to be necessary for maximal production of both Dha and pyochelin, but it was not required for Dha release from PchE and could not replace the thioesterase function specified by the C-terminal domain of PchF. In vitro, 2-aminobutyrate, a cysteine analog, was adenylated by purified PchE and PchF proteins. In vivo, this analog strongly interfered with Dha and pyochelin formation in a pchC deletion mutant but affected production of these metabolites only slightly in the wild type. Exogenously supplied cysteine overcame the negative effect of a pchC mutation to a large extent, whereas addition of salicylate did not. These data are in agreement with a role for PchC as an editing enzyme that removes wrongly charged molecules from the peptidyl carrier protein domains of PchE and PchF.
Figures
Similar articles
-
Assembly of the Pseudomonas aeruginosa nonribosomal peptide siderophore pyochelin: In vitro reconstitution of aryl-4, 2-bisthiazoline synthetase activity from PchD, PchE, and PchF.Biochemistry. 1999 Nov 9;38(45):14941-54. doi: 10.1021/bi991787c. Biochemistry. 1999. PMID: 10555976
-
Dihydroaeruginoic acid synthetase and pyochelin synthetase, products of the pchEF genes, are induced by extracellular pyochelin in Pseudomonas aeruginosa.Microbiology (Reading). 1998 Nov;144 ( Pt 11):3135-3148. doi: 10.1099/00221287-144-11-3135. Microbiology (Reading). 1998. PMID: 9846750
-
Essential PchG-dependent reduction in pyochelin biosynthesis of Pseudomonas aeruginosa.J Bacteriol. 2001 Feb;183(3):813-20. doi: 10.1128/JB.183.3.813-820.2001. J Bacteriol. 2001. PMID: 11208777 Free PMC article.
-
Iron acquisition with the natural siderophore enantiomers pyochelin and enantio-pyochelin in Pseudomonas species.Biometals. 2011 Jun;24(3):513-22. doi: 10.1007/s10534-010-9399-9. Epub 2010 Dec 25. Biometals. 2011. PMID: 21188474 Review.
-
Nonribosomal peptides for iron acquisition: pyochelin biosynthesis as a case study.Curr Opin Struct Biol. 2018 Dec;53:1-11. doi: 10.1016/j.sbi.2018.01.015. Epub 2018 Feb 20. Curr Opin Struct Biol. 2018. PMID: 29455106 Free PMC article. Review.
Cited by
-
Siderophore-based iron acquisition and pathogen control.Microbiol Mol Biol Rev. 2007 Sep;71(3):413-51. doi: 10.1128/MMBR.00012-07. Microbiol Mol Biol Rev. 2007. PMID: 17804665 Free PMC article. Review.
-
A novel streptococcal integrative conjugative element involved in iron acquisition.Mol Microbiol. 2008 Dec;70(5):1274-92. doi: 10.1111/j.1365-2958.2008.06481.x. Mol Microbiol. 2008. PMID: 18990191 Free PMC article.
-
Massively parallel mutant selection identifies genetic determinants of Pseudomonas aeruginosa colonization of Drosophila melanogaster.bioRxiv [Preprint]. 2023 Nov 21:2023.11.20.567573. doi: 10.1101/2023.11.20.567573. bioRxiv. 2023. Update in: mSystems. 2024 Mar 19;9(3):e0131723. doi: 10.1128/msystems.01317-23. PMID: 38045230 Free PMC article. Updated. Preprint.
-
In vitro kinetic analysis of substrate specificity in enterobactin biosynthetic lower pathway enzymes provides insight into the biochemical function of the hot dog-fold thioesterase EntH.Biochemistry. 2009 Jan 27;48(3):511-3. doi: 10.1021/bi802207t. Biochemistry. 2009. PMID: 19119850 Free PMC article.
-
Two new siderophores produced by Pseudomonas sp. NCIMB 10586: The anti-oomycete non-ribosomal peptide synthetase-dependent mupirochelin and the NRPS-independent triabactin.Front Microbiol. 2023 Mar 24;14:1143861. doi: 10.3389/fmicb.2023.1143861. eCollection 2023. Front Microbiol. 2023. PMID: 37032897 Free PMC article.
References
-
- Audenaert, K., T. Pattery, P. Cornelis, and M. Höfte. 2002. Induction of systemic resistance to Botrytis cinerea in tomato by Pseudomonas aeruginosa 7NSK2: role of salicylic acid, pyochelin, and pyocyanin. Mol. Plant-Microbe Interact. 15:1147-1156. - PubMed
-
- Butler, A. R., N. Bate, and E. Cundliffe. 1999. Impact of thioesterase activity on tylosin biosynthesis in Streptomyces fradiae. Chem. Biol. 6:287-292. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
