

Conserved tertiary base pairing ensures proper RNA folding and efficient assembly of the signal recognition particle Alu domain
- PMID: 15383645
- PMCID: PMC519120
- DOI: 10.1093/nar/gkh837
Conserved tertiary base pairing ensures proper RNA folding and efficient assembly of the signal recognition particle Alu domain
Abstract
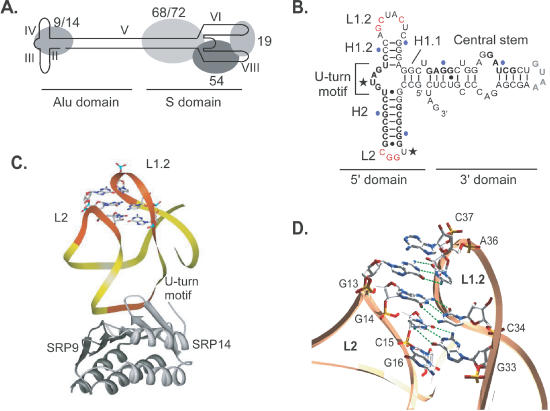
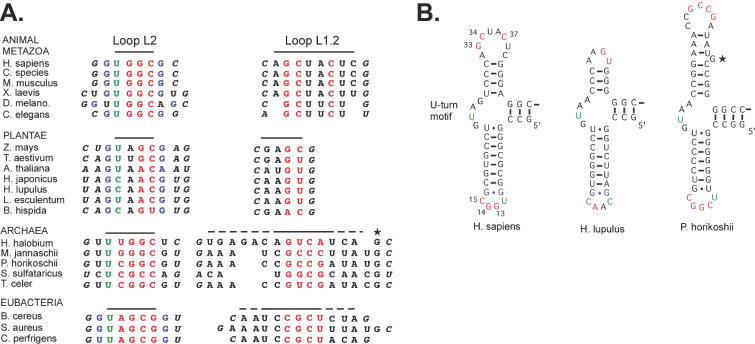
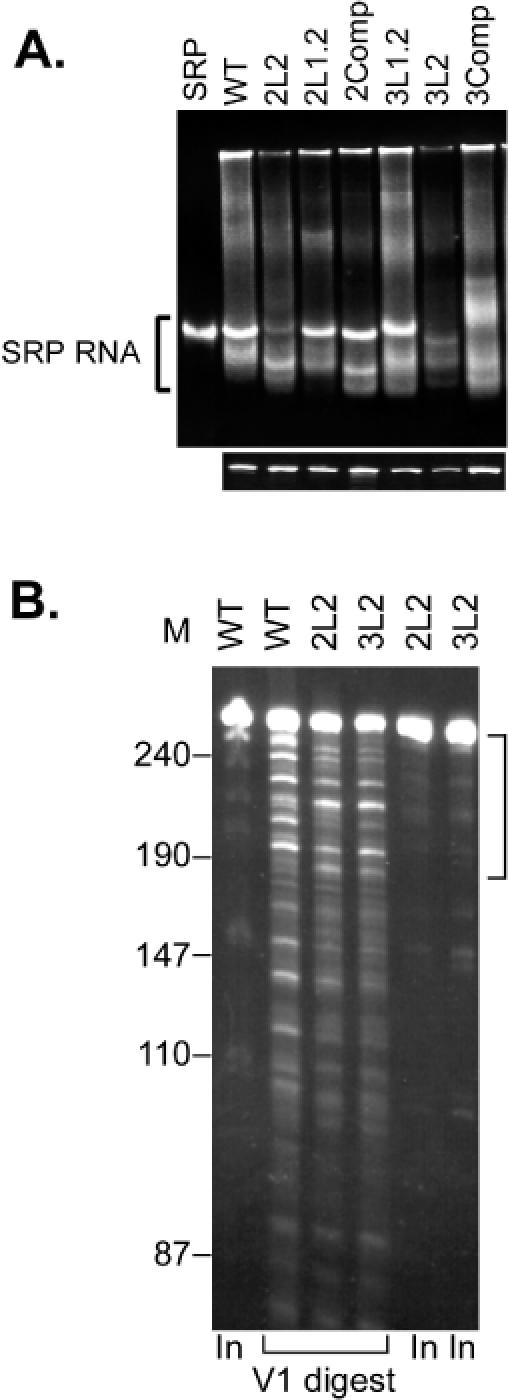
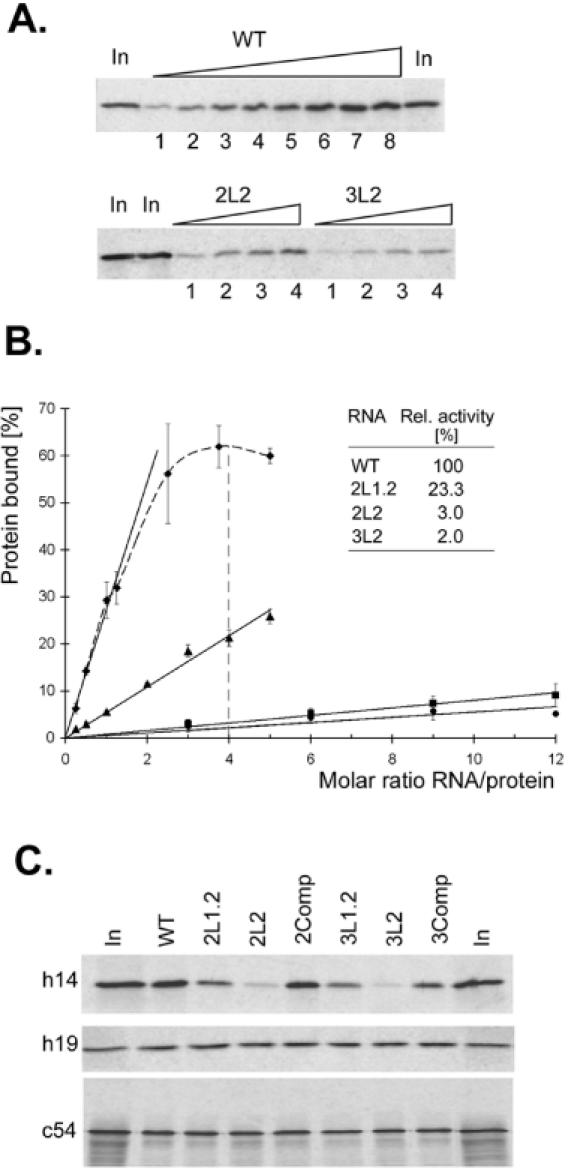
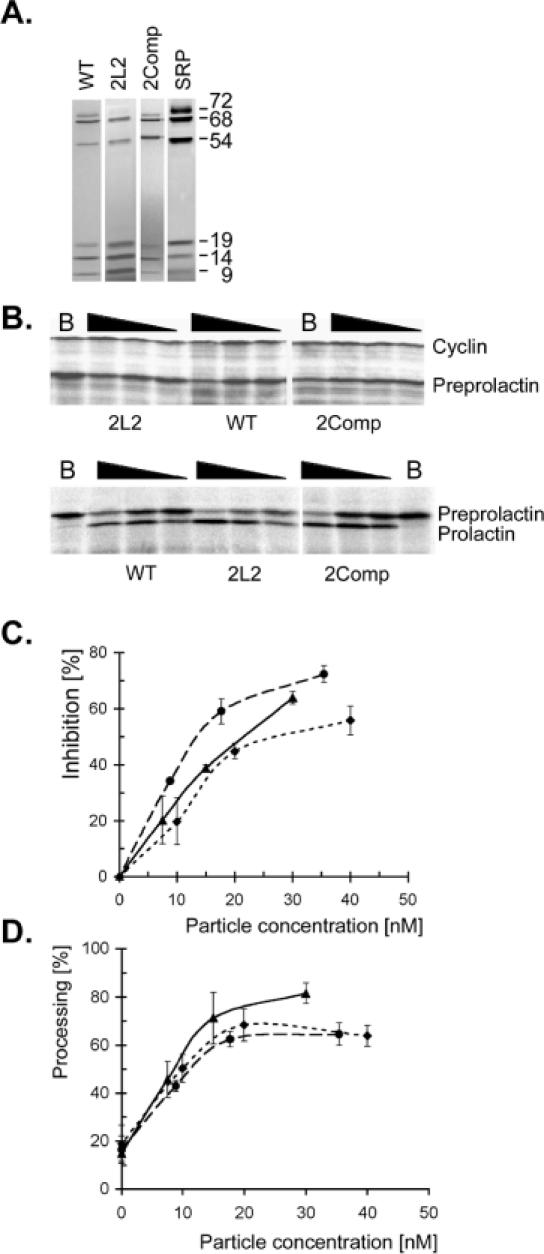
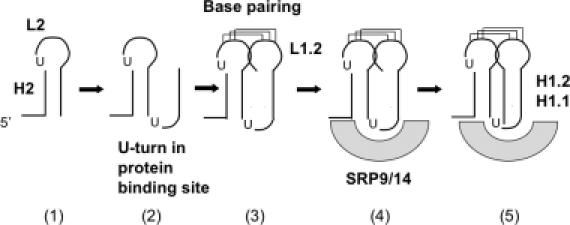
Proper folding of the RNA is an essential step in the assembly of functional ribonucleoprotein complexes. We examined the role of conserved base pairs formed between two distant loops in the Alu portion of the mammalian signal recognition particle RNA (SRP RNA) in SRP assembly and functions. Mutations disrupting base pairing interfere with folding of the Alu portion of the SRP RNA as monitored by probing the RNA structure and the binding of the protein SRP9/14. Complementary mutations rescue the defect establishing a role of the tertiary loop-loop interaction in RNA folding. The same mutations in the Alu domain have no major effect on binding of proteins to the S domain suggesting that the S domain can fold independently. Once assembled into a complete SRP, even particles that contain mutant RNA are active in arresting nascent chain elongation and translocation into microsomes, and, therefore, tertiary base pairing does not appear to be essential for these activities. Our results suggest a model in which the loop-loop interaction and binding of the protein SRP9/14 play an important role in the early steps of SRP RNA folding and assembly.
Figures
References
-
- Weeks K.M. (1997) Protein-facilitated RNA folding. Curr. Opin. Struct. Biol., 7, 336–342. - PubMed
-
- Walter P. and Johnson,A.E. (1994) Signal sequence recognition and protein targeting to the endoplasmic reticulum membrane. Annu. Rev. Cell Biol., 10, 87–119. - PubMed
-
- Johnson A.E. and van Waes,M.A. (1999) The translocon: a dynamic gateway at the ER membrane. Annu. Rev. Cell Dev. Biol., 15, 799–842. - PubMed
-
- Keenan R.J., Freymann,D.M., Stroud,R.M. and Walter,P. (2001) The signal recognition particle. Annu. Rev. Biochem., 70, 755–775. - PubMed
