

Requirements of fission yeast septins for complex formation, localization, and function
- PMID: 15385632
- PMCID: PMC532033
- DOI: 10.1091/mbc.e04-07-0640
Requirements of fission yeast septins for complex formation, localization, and function
Abstract
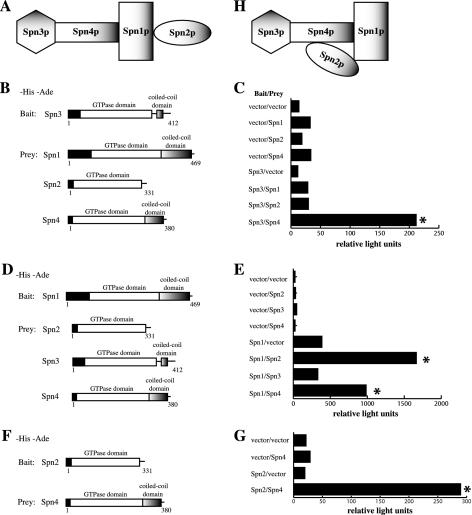
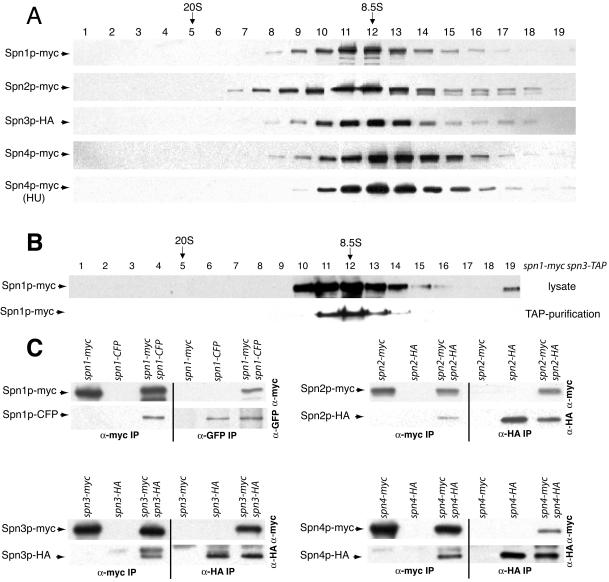
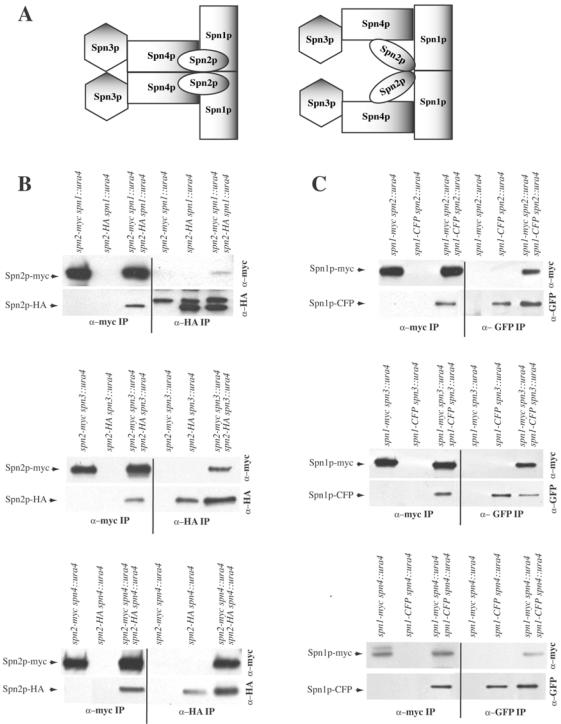
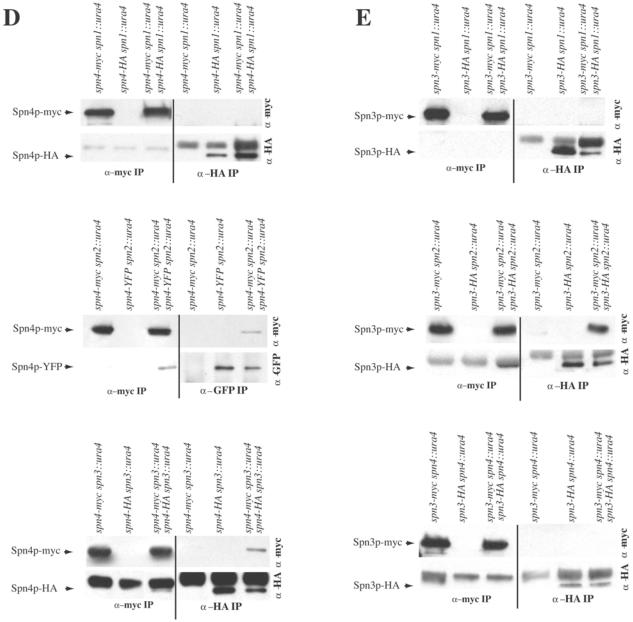
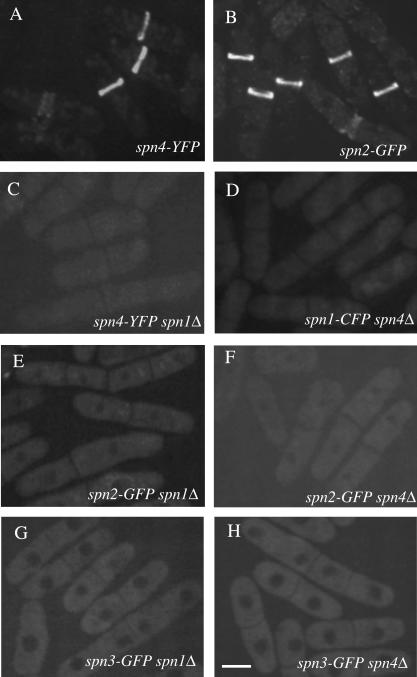
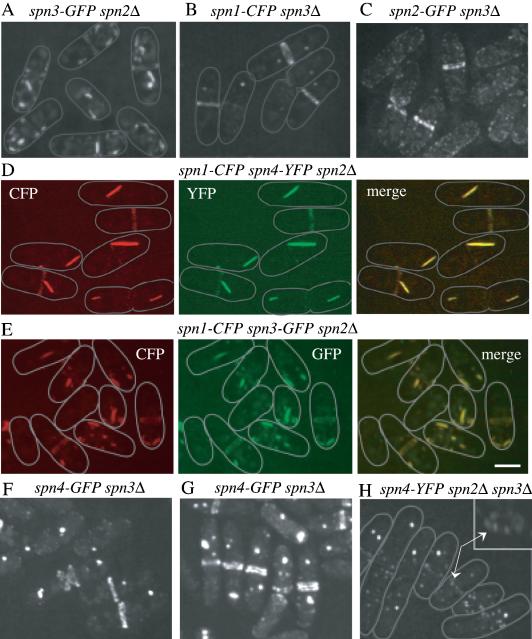
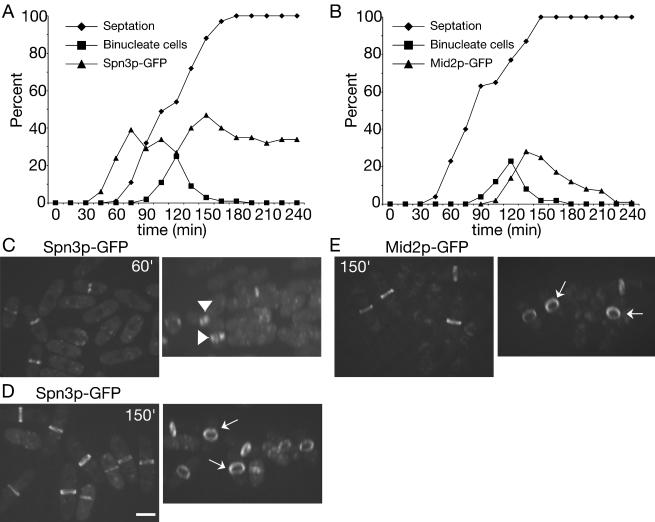
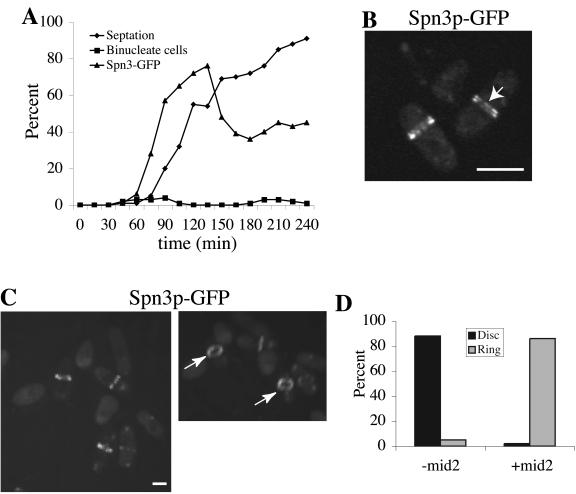
Septins are GTP binding proteins important for cytokinesis in many eukaryotes. The Schizosaccaromyces pombe genome sequence predicts orthologues of four of five Saccharomyces cerevisiae septins involved in cytokinesis and these are named Spns1-4p. That spns1-4 are not essential genes permitted the application of a combined genetic and proteomics approach to determine their functional relationships. Our findings indicate that Spns1-4p are present throughout interphase as a diffusely localized approximately 8.5S complex containing two copies of each septin linked together as a chain in the order Spn3p-Spn4p-Spn1p-Spn2p. Septin recruitment to the medial region of the cell is genetically separable from ring formation, and whereas it is normally restricted to mitosis, it can be promoted without activation of the mitotic cell cycle machinery. Coalescence into ring structures requires Spn1p and Spn4p associate with at least one other septin subunit and the expression of Mid2p that is normally restricted to mitosis. This study establishes the functional requirements for septin complex organization in vivo.
Figures

References
-
- Bahler, J., Wu, J.Q., Longtine, M.S., Shah, N.G., McKenzie, A., 3rd, Steever, A.B., Wach, A., Philippsen, P., and Pringle, J.R. (1998). Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14, 943-951. - PubMed
-
- Burns, C.G., Ohi, R., Mehta, S., O'Toole, E.T., Winey, M., Clark, T.A., Sugnet, C.W., Ares, M., Jr., and Gould, K.L. (2002). Removal of a single alpha-tubulin gene intron suppresses cell cycle arrest phenotypes of splicing factor mutations in Saccharomyces cerevisiae. Mol. Cell. Biol. 22, 801-815. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
