

Structural and functional anatomy of the globular domain of complement protein C1q
- PMID: 15388251
- PMCID: PMC3818097
- DOI: 10.1016/j.imlet.2004.06.015
Structural and functional anatomy of the globular domain of complement protein C1q
Erratum in
- Immunol Lett. 2005 Oct 15;101(1):116
Abstract
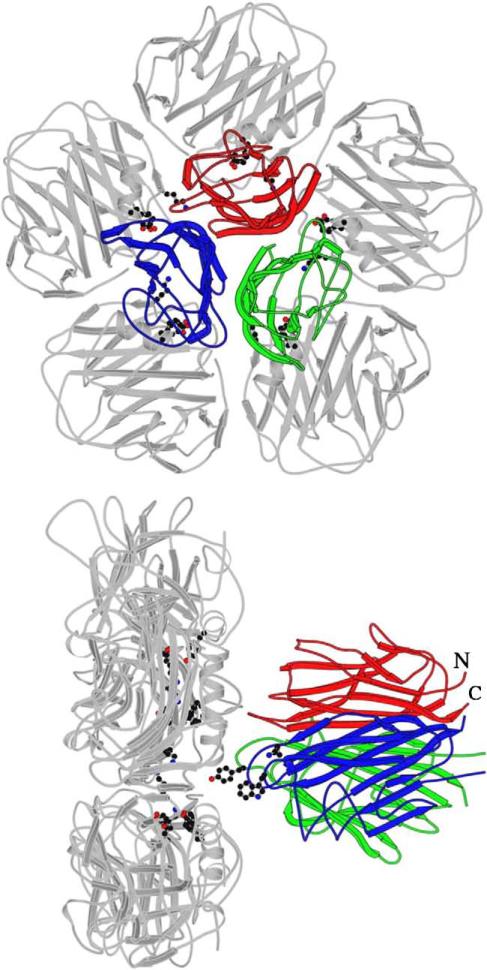
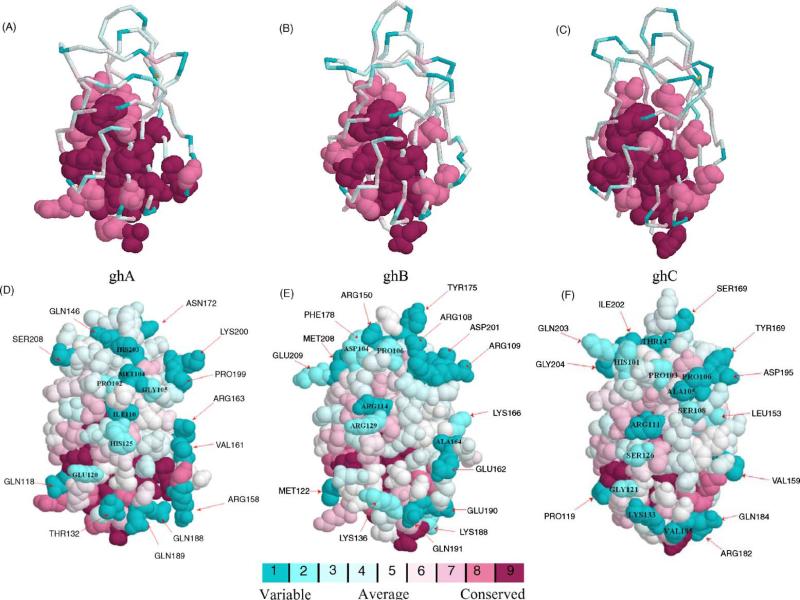
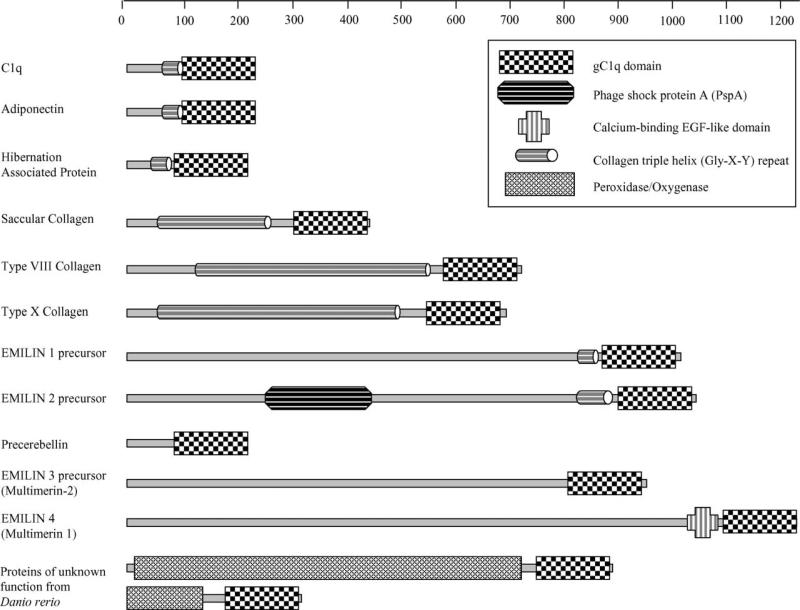
C1q is the first subcomponent of the classical pathway of the complement system and a major connecting link between innate and acquired immunity. As a versatile charge pattern recognition molecule, C1q is capable of engaging a broad range of ligands via its heterotrimeric globular domain (gC1q) which is composed of the C-terminal regions of its A (ghA), B (ghB) and C (ghC) chains. Recent studies using recombinant forms of ghA, ghB and ghC have suggested that the gC1q domain has a modular organization and each chain can have differential ligand specificity. The crystal structure of the gC1q, molecular modeling and protein engineering studies have combined to illustrate how modular organization, charge distribution and the spatial orientation of the heterotrimeric assembly offer versatility of ligand recognition to C1q. Although the biochemical and structural studies have provided novel insights into the structure-function relationships within the gC1q domain, they have also raised many unexpected issues for debate.
Figures
Similar articles
-
Localization of ligand-binding sites on human C1q globular head region using recombinant globular head fragments and single-chain antibodies.Biochim Biophys Acta. 2003 Nov 3;1652(1):64-74. doi: 10.1016/j.bbapap.2003.08.003. Biochim Biophys Acta. 2003. PMID: 14580997
-
Interaction of C1q with IgG1, C-reactive protein and pentraxin 3: mutational studies using recombinant globular head modules of human C1q A, B, and C chains.Biochemistry. 2006 Apr 4;45(13):4093-104. doi: 10.1021/bi052646f. Biochemistry. 2006. PMID: 16566583 Free PMC article.
-
Existence of different but overlapping IgG- and IgM-binding sites on the globular domain of human C1q.Biochemistry. 2006 Aug 22;45(33):9979-88. doi: 10.1021/bi060539v. Biochemistry. 2006. PMID: 16906756
-
C1q and its growing family.Immunobiology. 2007;212(4-5):253-66. doi: 10.1016/j.imbio.2006.11.001. Epub 2006 Dec 12. Immunobiology. 2007. PMID: 17544811 Review.
-
C1q and tumor necrosis factor superfamily: modularity and versatility.Trends Immunol. 2004 Oct;25(10):551-61. doi: 10.1016/j.it.2004.08.006. Trends Immunol. 2004. PMID: 15364058 Review.
Cited by
-
Complement System Part I - Molecular Mechanisms of Activation and Regulation.Front Immunol. 2015 Jun 2;6:262. doi: 10.3389/fimmu.2015.00262. eCollection 2015. Front Immunol. 2015. PMID: 26082779 Free PMC article. Review.
-
A C1q domain containing protein from scallop Chlamys farreri serving as pattern recognition receptor with heat-aggregated IgG binding activity.PLoS One. 2012;7(8):e43289. doi: 10.1371/journal.pone.0043289. Epub 2012 Aug 15. PLoS One. 2012. PMID: 22905248 Free PMC article.
-
Microglia-Astrocyte Communication via C1q Contributes to Orofacial Neuropathic Pain Associated with Infraorbital Nerve Injury.Int J Mol Sci. 2020 Sep 17;21(18):6834. doi: 10.3390/ijms21186834. Int J Mol Sci. 2020. PMID: 32957694 Free PMC article.
-
Cryo-Electron Microscopy and Biochemical Analysis Offer Insights Into the Effects of Acidic pH, Such as Occur During Acidosis, on the Complement Binding Properties of C-Reactive Protein.Front Immunol. 2021 Dec 16;12:757633. doi: 10.3389/fimmu.2021.757633. eCollection 2021. Front Immunol. 2021. PMID: 34975846 Free PMC article.
-
Comparative genomic analysis of eutherian adiponectin genes.Heliyon. 2018 Jun 6;4(6):e00647. doi: 10.1016/j.heliyon.2018.e00647. eCollection 2018 Jun. Heliyon. 2018. PMID: 30003153 Free PMC article.
References
-
- Arlaud GJ, Gaboriaud C, Thielens NM, Rossi V, Bersch B, Hernandez JF, Fontecilla-Camps JC. Structural biology of C1: dissection of a complex molecular machinery. Immunol Rev. 2001;180:136–45. - PubMed
-
- Arlaud GJ, Gaboriaud C, Thielens NM, Budayova-Spano M, Rossi V, Fontecilla-Camps JC. Structural biology of the C1 complex of complement unveils the mechanisms of its activation and proteolytic activity. Mol Immunol. 2002;39:383–94. - PubMed
-
- Arlaud GJ, Gaboriaud C, Thielens NM, Rossi V. Structural biology of C1. Biochem Soc Trans. 2002;30:1001–6. - PubMed
-
- Gaboriaud C, Juanhuix J, Gruez A, Lacroix M, Darnault C, Pignol D, Verger D, Fontecilla-Camps JC, Arlaud GJ. The crystal structure of the globular head of complement protein C1q provides a basis for its versatile recognition properties. J Biol Chem. 2003;278:46974–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
