

Innate immune response of corneal epithelial cells to Staphylococcus aureus infection: role of peptidoglycan in stimulating proinflammatory cytokine secretion
- PMID: 15452057
- PMCID: PMC2666393
- DOI: 10.1167/iovs.04-0467
Innate immune response of corneal epithelial cells to Staphylococcus aureus infection: role of peptidoglycan in stimulating proinflammatory cytokine secretion
Abstract
Purpose: This study sought to elucidate the innate immune responses of cultured human corneal epithelial cells (HCECs) to infection by the Gram-positive bacterium Staphylococcus aureus and to determine the underlying mechanisms.
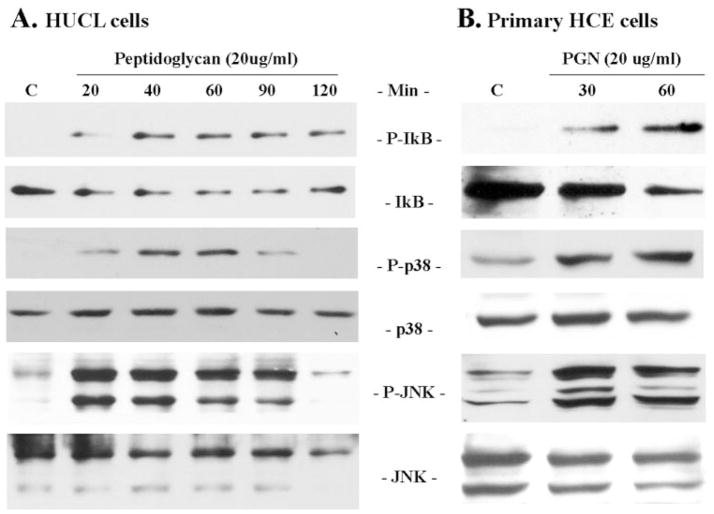
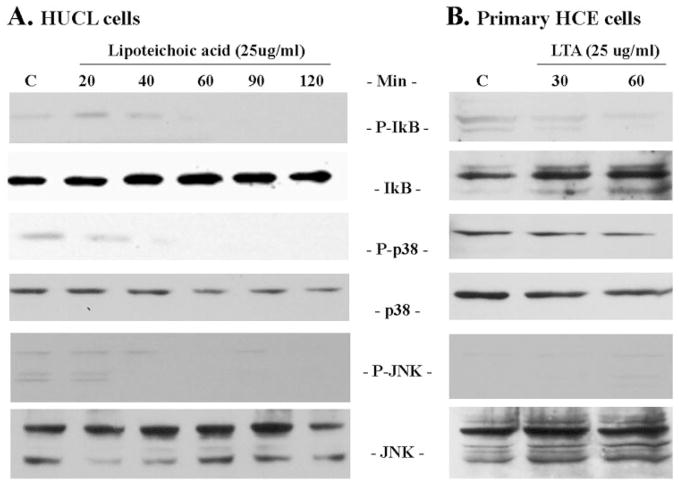
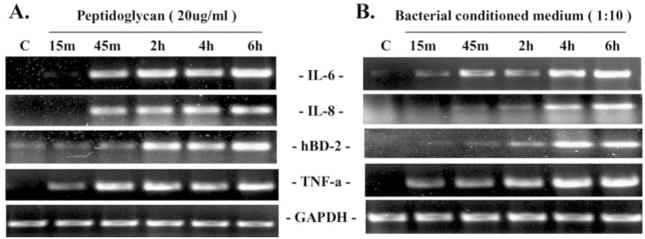
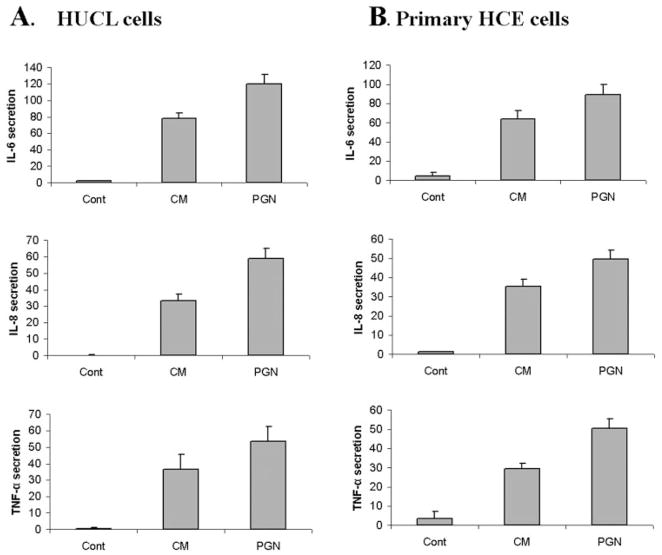
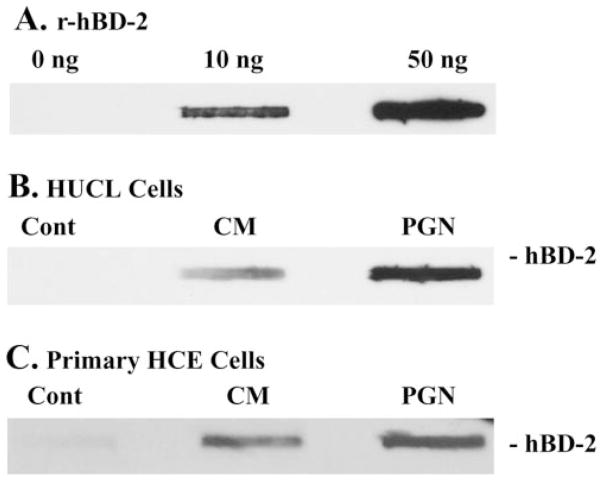
Methods: HUCL, a telomerase-immortalized HCEC line, and primary cultures of HCECs were challenged with live or heat-killed S. aureus, its exoproducts, or cell wall components lipoteichoic acid (LTA) and peptidoglycan (PGN). IkappaB-alpha phosphorylation and degradation as well as phosphorylation of MAPKs, p38, and JNK-1/2, were assessed by Western blot analysis. The expression of interleukin (IL)-6, IL-8, TNF-alpha, and beta-defensin-2 were determined using RT-PCR and secretion of IL-6, IL-8, TNF-alpha, and beta-defensin were measured using enzyme-linked immunosorbent assay and immunoblot analysis of culture medium.
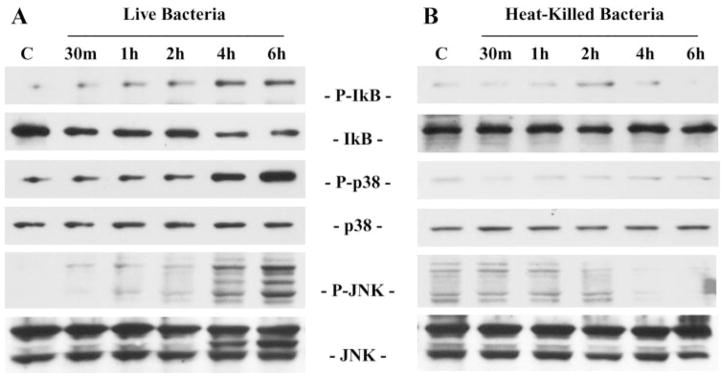
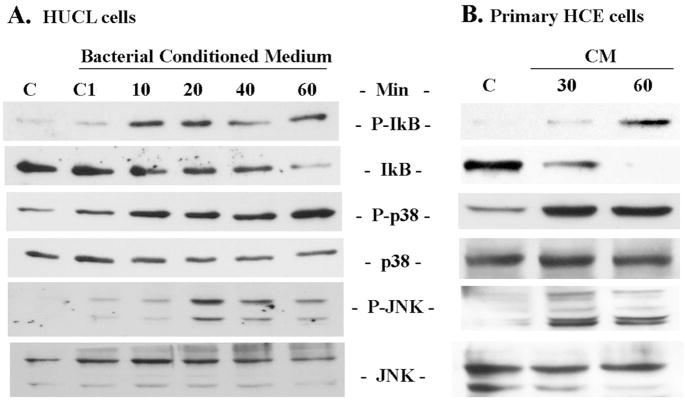
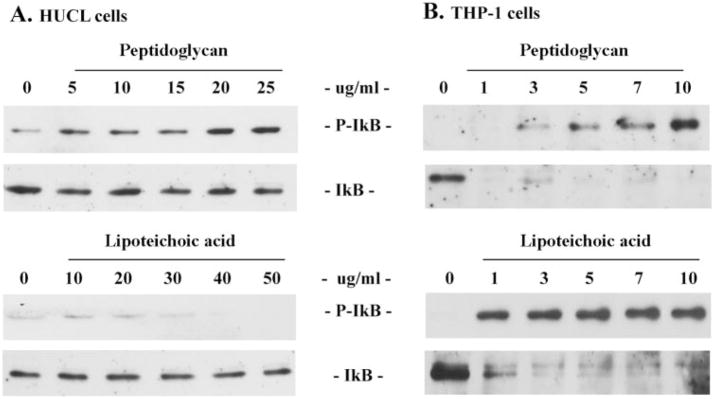
Results: Exposure of HUCL cells to live, but not heat-killed, S. aureus resulted in NF-kappaB activation in a time-dependent manner, as assessed by the increase in IkappaB-alpha phosphorylation and degradation. Live bacteria also activated the p38 and JNK pathways. The effects of live bacteria on HUCL cells may be attributable to bacterial exoproducts, since the conditioned medium of S. aureus also effectively stimulated these signaling pathways. PGN, but not LTA, activated the NF-kappaB and MAPK pathways in a dose- and time-dependent manner. Concomitant with activation of NF-kappaB and MAPKs, transcriptional expression of IL-6, IL-8, TNF-alpha, and beta-defensin-2 were induced in cells challenged with bacterial exoproducts and PGN. Secretion of IL-6, IL-8, TNF-alpha, and beta-defensin-2 were also significantly increased in HCECs in response to bacterial exoproducts and PGN challenge.
Conclusions: Corneal epithelial cells possess the ability to recognize the presence of Gram-positive bacteria and to initiate the innate immune responses by the expression and/or release of proinflammatory cytokines and beta-defensin-2 in the cornea.
Figures
Similar articles
-
Inflammatory responses of corneal epithelial cells to Pseudomonas aeruginosa infection.Curr Eye Res. 2005 Jul;30(7):527-34. doi: 10.1080/02713680590968150. Curr Eye Res. 2005. PMID: 16020286 Free PMC article.
-
Modulation of corneal epithelial innate immune response to pseudomonas infection by flagellin pretreatment.Invest Ophthalmol Vis Sci. 2007 Oct;48(10):4664-70. doi: 10.1167/iovs.07-0473. Invest Ophthalmol Vis Sci. 2007. PMID: 17898290 Free PMC article.
-
Toll-like receptor 2-mediated expression of beta-defensin-2 in human corneal epithelial cells.Microbes Infect. 2006 Feb;8(2):380-9. doi: 10.1016/j.micinf.2005.07.006. Epub 2005 Sep 15. Microbes Infect. 2006. PMID: 16242370 Free PMC article.
-
Defensin expression by the cornea: multiple signalling pathways mediate IL-1beta stimulation of hBD-2 expression by human corneal epithelial cells.Invest Ophthalmol Vis Sci. 2003 May;44(5):1859-65. doi: 10.1167/iovs.02-0787. Invest Ophthalmol Vis Sci. 2003. PMID: 12714616 Free PMC article.
-
Toll-like receptor 5-mediated corneal epithelial inflammatory responses to Pseudomonas aeruginosa flagellin.Invest Ophthalmol Vis Sci. 2003 Oct;44(10):4247-54. doi: 10.1167/iovs.03-0219. Invest Ophthalmol Vis Sci. 2003. PMID: 14507868
Cited by
-
Human conjunctival epithelial cells lack lipopolysaccharide responsiveness due to deficient expression of MD2 but respond after interferon-gamma priming or soluble MD2 supplementation.Inflammation. 2005 Dec;29(4-6):170-81. doi: 10.1007/s10753-006-9014-y. Inflammation. 2005. PMID: 17093906
-
Sodium Octanoate Modulates the Innate Immune Response of Bovine Mammary Epithelial Cells through the TLR2/P38/JNK/ERK1/2 Pathway: Implications during Staphylococcus aureus Internalization.Front Cell Infect Microbiol. 2017 Mar 15;7:78. doi: 10.3389/fcimb.2017.00078. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28361042 Free PMC article.
-
Myxinidin2 and myxinidin3 suppress inflammatory responses through STAT3 and MAPKs to promote wound healing.Oncotarget. 2017 Sep 15;8(50):87582-87597. doi: 10.18632/oncotarget.20908. eCollection 2017 Oct 20. Oncotarget. 2017. PMID: 29152103 Free PMC article.
-
Staphylococcus aureus protein A induced inflammatory response in human corneal epithelial cells.Biochem Biophys Res Commun. 2007 Mar 23;354(4):955-61. doi: 10.1016/j.bbrc.2007.01.072. Epub 2007 Jan 23. Biochem Biophys Res Commun. 2007. PMID: 17270147 Free PMC article.
-
Expression of toll-like receptors in human limbal and conjunctival epithelial cells.Mol Vis. 2007 Jun 8;13:813-22. Mol Vis. 2007. PMID: 17615542 Free PMC article.
References
-
- Alexandrakis G, Alfonso EC, Miller D. Shifting trends in bacterial keratitis in south Florida and emerging resistance to fluoroquinolones. Ophthalmology. 2000;107:1497–1502. - PubMed
-
- Kurpakus-Wheater M, Kernacki KA, Hazlett LD. Maintaining corneal integrity how the “window” stays clear. Prog Histochem Cytochem. 2001;36:185–259. - PubMed
-
- Lyczak JB, Cannon CL, Pier GB. Establishment of Pseudomonas aeruginosa infection: lessons from a versatile opportunist. Microbes Infect. 2000;2:1051–1060. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
