

Coordinate expression and trans presentation of interleukin (IL)-15Ralpha and IL-15 supports natural killer cell and memory CD8+ T cell homeostasis
- PMID: 15452177
- PMCID: PMC2213280
- DOI: 10.1084/jem.20041389
Coordinate expression and trans presentation of interleukin (IL)-15Ralpha and IL-15 supports natural killer cell and memory CD8+ T cell homeostasis
Abstract
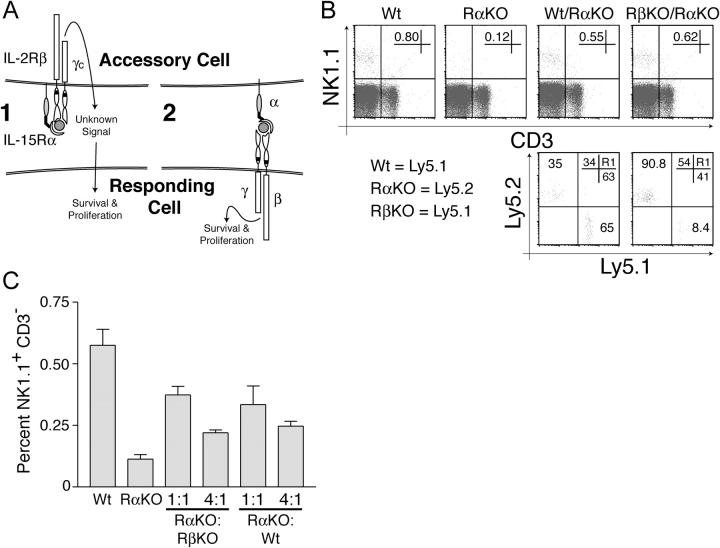
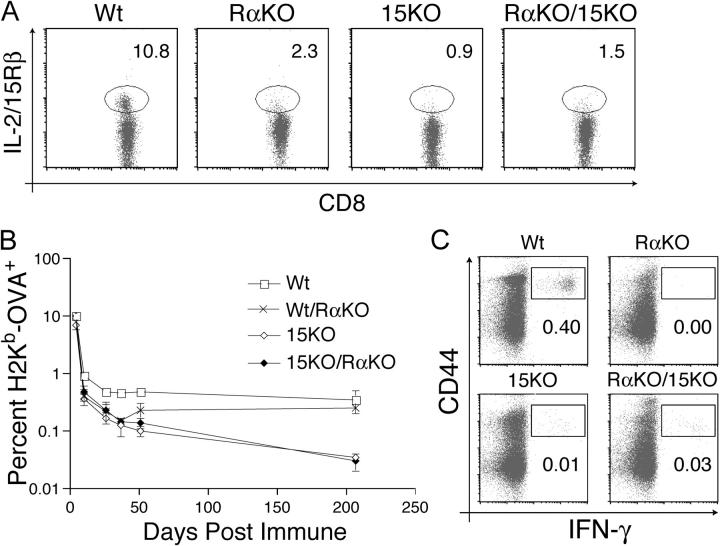
The high affinity interleukin (IL)-15 receptor, IL-15Ralpha, is essential for supporting lymphoid homeostasis. To assess whether IL-15Ralpha's role in vivo is to trans present IL-15, we generated mixed bone marrow chimera from IL-15Ralpha- and IL-2/15Rbeta-deficient mice. We find that IL-15Ralpha-competent, IL-2/15Rbeta-deficient cells are able to support IL-15Ralpha-deficient natural killer (NK) and memory CD8+ T cells, thus ruling out secondary signals on these cells and demonstrating that IL-15Ralpha-mediated presentation of IL-15 in trans is the primary mechanism by which IL-15Ralpha functions in vivo. Surprisingly, using IL-15- and IL-15Ralpha-deficient mixed chimera, we also find that IL-15 and IL-15Ralpha must be expressed by the same cells to present IL-15 in trans, indicating that IL-15Ralpha is required on a cellular level for the elaboration of IL-15. These studies indicate that IL-15Ralpha defines homeostatic niches for NK and memory CD8+ T cells by controlling both the production and the presentation of IL-15 in trans to NK and CD8+ memory T cells.
Figures




References
-
- Schluns, K.S., W.C. Kieper, S.C. Jameson, and L. Lefrancois. 2000. Interleukin-7 mediates the homeostasis of naive and memory CD8 T cells in vivo. Nat. Immunol. 1:426–432. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
