

Transmembrane peptide-induced lipid sorting and mechanism of Lalpha-to-inverted phase transition using coarse-grain molecular dynamics
- PMID: 15454415
- PMCID: PMC1304638
- DOI: 10.1529/biophysj.104.040311
Transmembrane peptide-induced lipid sorting and mechanism of Lalpha-to-inverted phase transition using coarse-grain molecular dynamics
Abstract
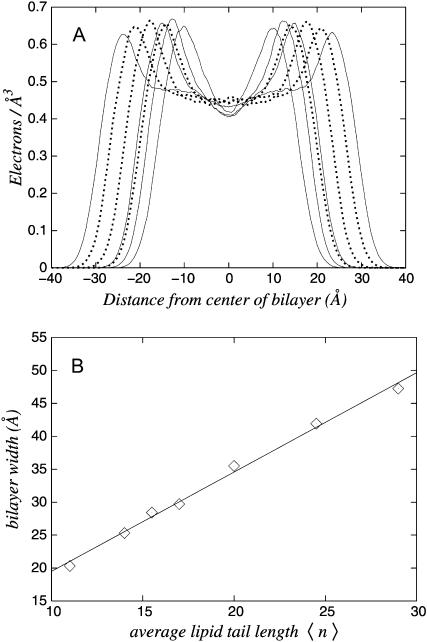
Molecular dynamics results are presented for a coarse-grain model of 1,2-di-n-alkanoyl-sn-glycero-3-phosphocholine, water, and a capped cylindrical model of a transmembrane peptide. We first demonstrate that different alkanoyl-length lipids are miscible in the liquid-disordered lamellar (Lalpha) phase. The transmembrane peptide is constructed of hydrophobic sites with hydrophilic caps. The hydrophobic length of the peptide is smaller than the hydrophobic thickness of a bilayer consisting of an equal mixture of long and short alkanoyl tail lipids. When incorporated into the membrane, a meniscus forms in the vicinity of the peptide and the surrounding area is enriched in the short lipid. The meniscus region draws water into it. In the regions that are depleted of water, the bilayers can fuse. The lipid headgroups then rearrange to solvate the newly formed water pores, resulting in an inverted phase. This mechanism appears to be a viable pathway for the experimentally observed Lalpha-to-inverse hexagonal (HII) peptide-induced phase transition.
Copyright 2004 Biophysical Society
Figures








References
-
- Betanzos, M., C. S. Chiang, H. R. Guy, and S. Sukharev. 2002. A large iris-like expansion of a mechanosensitive channel protein induced by membrane tension. Nat. Struct. Biol. 9:704–710. - PubMed
-
- Caffrey, M. 2000. A lipid's eye view of membrane protein crystallization in mesophases. Curr. Opin. Struct. Biol. 10:486–497. - PubMed
-
- Dan, N., P. Pincus, and S. A. Safran. 1993. Membrane-induced interactions between inclusions. Langmuir. 9:2768–2771.
-
- de Planque, M. R. R., E. Goormaghtigh, D. V. Greathouse, R. E. Koeppe II, J. A. W. Kruijtzer, R. M. J. Liskamp, B. de Kruijff, and J. A. Killian. 2001. Sensitivity of single membrane-spanning α-helical peptides to hydrophobic mismatch with a lipid bilayer: effects on backbone structure, orientation, and extent of membrane incorporation. Biochemistry. 40:5000–5010. - PubMed
-
- de Planque, M. R. R., D. V. Greathouse, R. E. Koeppe II, H. Schäfer, D. Marsh, and J. A. Killian. 1998. Influence of lipid/peptide hydrophobic mismatch on the thickness of diacylphosphatidylcholine bilayers. A 2H NMR and ESR study using designed transmembrane α-helical peptides and gramicidin A. Biochemistry. 37:9333–9345. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
