

RNA polymerase II carboxy-terminal domain phosphorylation is required for cotranscriptional pre-mRNA splicing and 3'-end formation
- PMID: 15456870
- PMCID: PMC517882
- DOI: 10.1128/MCB.24.20.8963-8969.2004
RNA polymerase II carboxy-terminal domain phosphorylation is required for cotranscriptional pre-mRNA splicing and 3'-end formation
Abstract
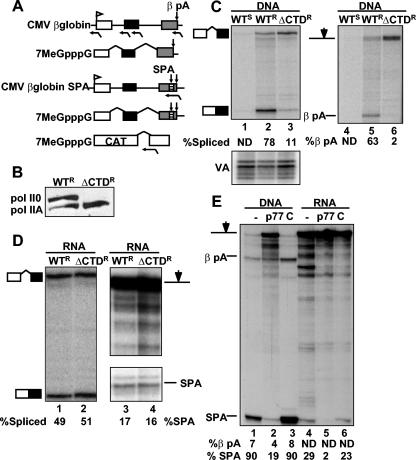
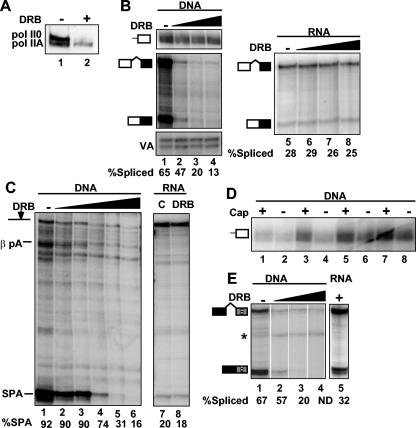
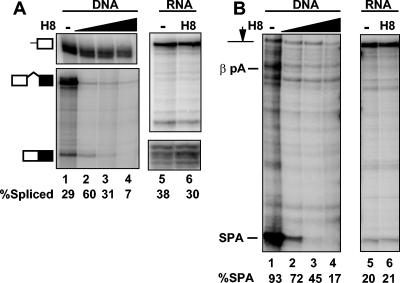
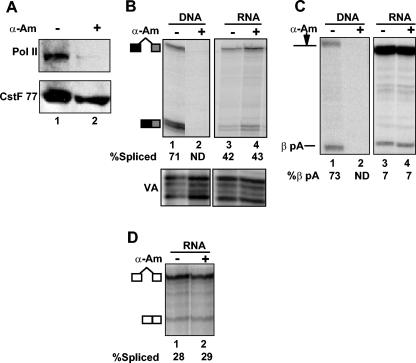
We investigated the role of RNA polymerase II (pol II) carboxy-terminal domain (CTD) phosphorylation in pre-mRNA processing coupled and uncoupled from transcription in Xenopus oocytes. Inhibition of CTD phosphorylation by the kinase inhibitors 5,6-dichloro-1beta-D-ribofuranosyl-benzimidazole and H8 blocked transcription-coupled splicing and poly(A) site cleavage. These experiments suggest that pol II CTD phosphorylation is required for efficient pre-mRNA splicing and 3'-end formation in vivo. In contrast, processing of injected pre-mRNA was unaffected by either kinase inhibitors or alpha-amanitin-induced depletion of pol II. pol II therefore does not appear to participate directly in posttranscriptional processing, at least in frog oocytes. Together these experiments show that the influence of the phosphorylated CTD on pre-mRNA splicing and 3'-end processing is mediated by transcriptional coupling.
Figures
References
-
- Ahn, S. H., M. Kim, and S. Buratowski. 2004. Phosphorylation of serine 2 within the rna polymerase II C-terminal domain couples transcription and 3′ end processing. Mol. Cell. 13:67-76. - PubMed
-
- Bauren, G., and L. Wieslander. 1994. Splicing of Balbiani ring 1 gene pre-mRNA occurs simultaneously with transcription. Cell 76:183-192. - PubMed
-
- Bentley, D. 2002. The mRNA assembly line: transcription and processing machines in the same factory. Curr. Opin. Cell Biol. 14:336-342. - PubMed
-
- Beyer, A. L., and Y. N. Osheim. 1988. Splice site selection, rate of splicing, and alternative splicing on nascent transcripts. Genes Dev. 2:754-765. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources