

PHR1, a PH domain-containing protein expressed in primary sensory neurons
- PMID: 15456885
- PMCID: PMC517893
- DOI: 10.1128/MCB.24.20.9137-9151.2004
PHR1, a PH domain-containing protein expressed in primary sensory neurons
Abstract
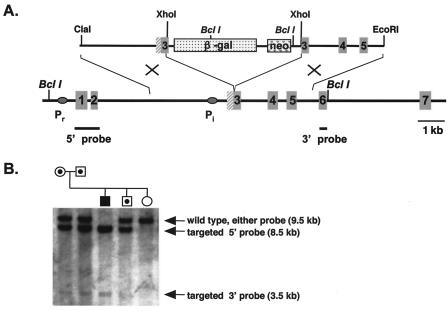
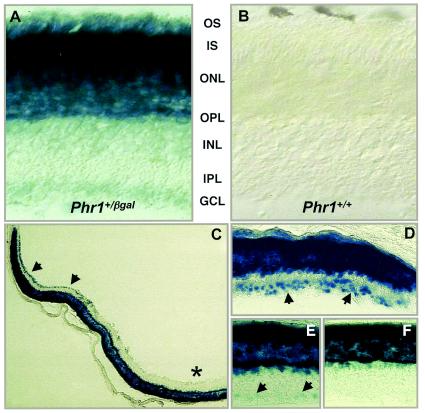
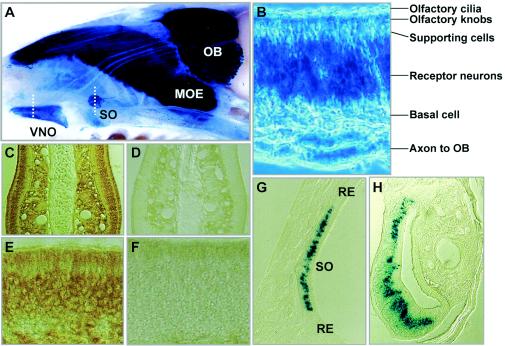
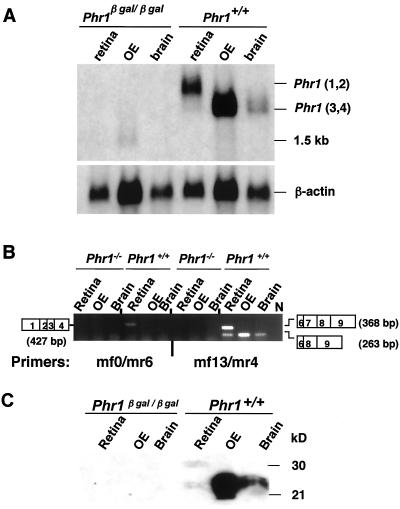
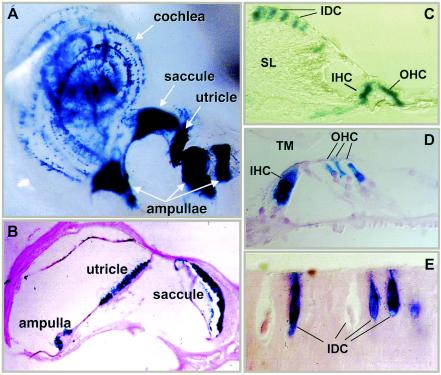
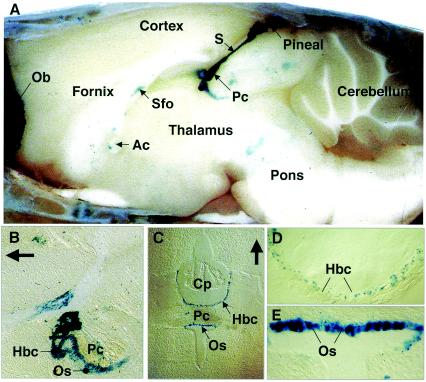
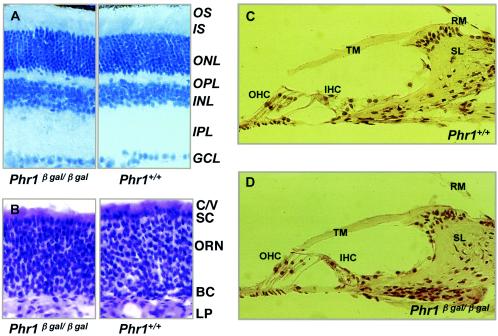
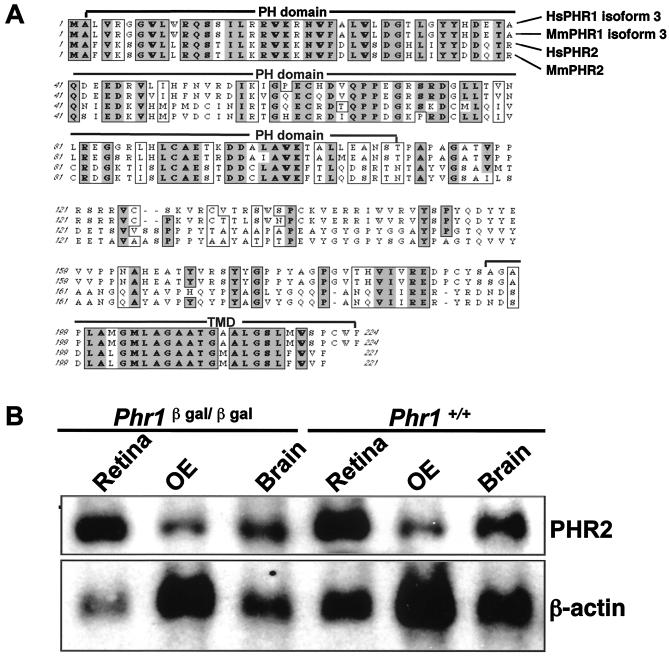
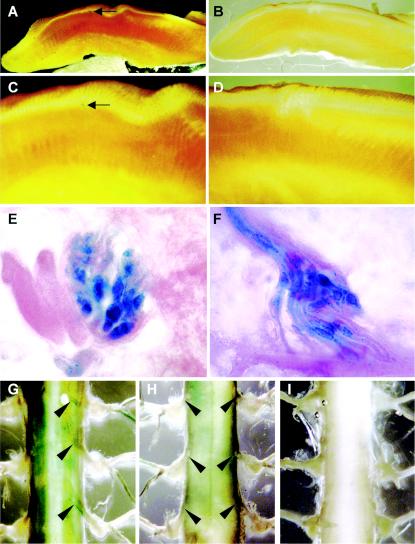
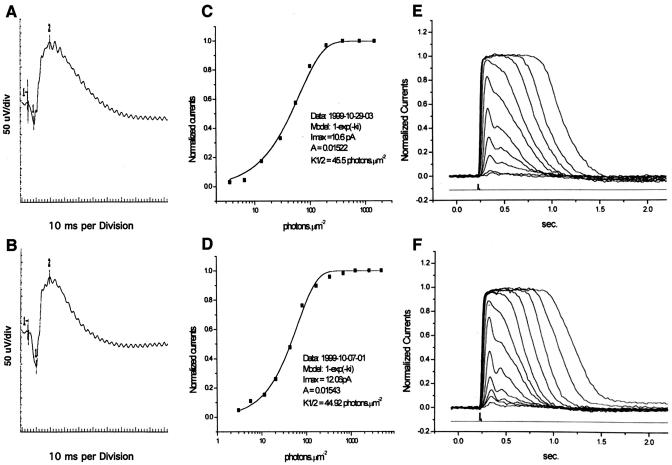
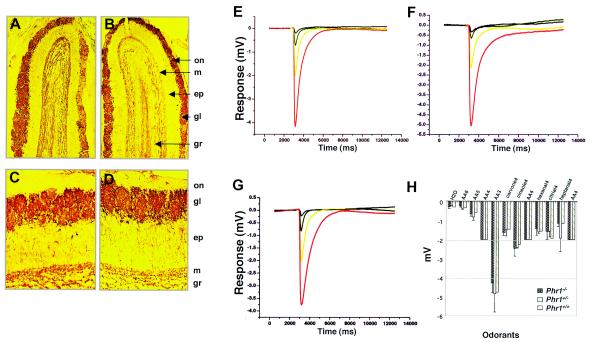
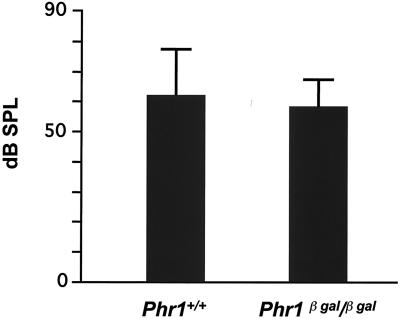
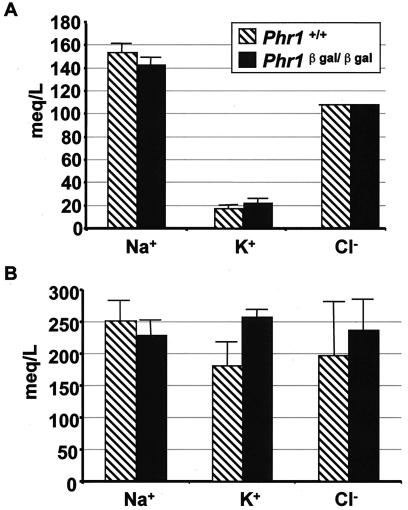
Previously, we identified PHR1 as an abundantly expressed gene in photoreceptors and showed that it encodes four isoforms, each with N-terminal pleckstrin homology (PH) and C-terminal transmembrane domains. To better understand PHR1 function and expression, we made a Phr1 null mouse by inserting a beta-galactosidase/neor cassette into exon 3. In addition to photoreceptors, we found abundant expression of specific Phr1 splice forms in olfactory receptor neurons and vestibular and cochlear hair cells. We also found Phr1 expression in cells with a possible sensory function, including peripheral retinal ganglion cells, cochlear interdental cells, and neurons of the circumventricular organ. Despite this discrete expression in known and putative sensory neurons, mice lacking PHR1 do not have overt sensory deficits.
Figures
Similar articles
-
PHR1 encodes an abundant, pleckstrin homology domain-containing integral membrane protein in the photoreceptor outer segments.J Biol Chem. 1999 Dec 10;274(50):35676-85. doi: 10.1074/jbc.274.50.35676. J Biol Chem. 1999. PMID: 10585447
-
PHR1 is a vesicle-bound protein abundantly expressed in mature olfactory neurons.Laryngoscope. 2010 May;120(5):1002-10. doi: 10.1002/lary.20779. Laryngoscope. 2010. PMID: 20301200
-
PHR1, an integral membrane protein of the inner ear sensory cells, directly interacts with myosin 1c and myosin VIIa.J Cell Sci. 2005 Jul 1;118(Pt 13):2891-9. doi: 10.1242/jcs.02424. J Cell Sci. 2005. PMID: 15976448
-
Neurod1 regulates survival and formation of connections in mouse ear and brain.Cell Tissue Res. 2010 Jul;341(1):95-110. doi: 10.1007/s00441-010-0984-6. Epub 2010 May 30. Cell Tissue Res. 2010. PMID: 20512592 Free PMC article.
-
Neurotrophins in the ear: their roles in sensory neuron survival and fiber guidance.Prog Brain Res. 2004;146:265-78. doi: 10.1016/S0079-6123(03)46017-2. Prog Brain Res. 2004. PMID: 14699969 Review.
Cited by
-
Inactivation of the miR-183/96/182 Cluster Decreases the Severity of Pseudomonas aeruginosa-Induced Keratitis.Invest Ophthalmol Vis Sci. 2016 Apr;57(4):1506-17. doi: 10.1167/iovs.16-19134. Invest Ophthalmol Vis Sci. 2016. PMID: 27035623 Free PMC article.
-
The single-cell transcriptomic landscape of the topological differences in mammalian auditory receptors.Sci China Life Sci. 2024 Nov;67(11):2398-2410. doi: 10.1007/s11427-024-2672-1. Epub 2024 Jul 29. Sci China Life Sci. 2024. PMID: 39083201
-
A novel locus for autosomal recessive nonsyndromic hearing impairment, DFNB63, maps to chromosome 11q13.2-q13.4.J Mol Med (Berl). 2007 Apr;85(4):397-404. doi: 10.1007/s00109-006-0136-3. Epub 2007 Jan 9. J Mol Med (Berl). 2007. PMID: 17211611
-
In vitro interaction between coxsackievirus B3 VP1 protein and human pleckstrin homology domain retinal protein (PHR1).Virus Genes. 2015 Oct;51(2):182-9. doi: 10.1007/s11262-015-1241-6. Epub 2015 Aug 30. Virus Genes. 2015. PMID: 26318175
-
Inactivation of the microRNA-183/96/182 cluster results in syndromic retinal degeneration.Proc Natl Acad Sci U S A. 2013 Feb 5;110(6):E507-16. doi: 10.1073/pnas.1212655110. Epub 2013 Jan 22. Proc Natl Acad Sci U S A. 2013. PMID: 23341629 Free PMC article.
References
-
- Akazawa, C., M. Ishibashi, C. Shimizu, S. Nakanshi, and R. Kageyama. 1995. A mammalian helix-loop-helix factor structurally related to the product of Drosophila proneural gene atonal is a positive transcriptional regulator expressed in the developing nervous system. J. Biol. Chem. 270:8730-8738. - PubMed
-
- Andrews, K., P. Potdar, P. Nettesheim, and L. E. Ostrowski. 2000. KPL1, which encodes a novel PH domain-containing protein, is induced during ciliated cell differentiation of rat tracheal epithelial cells. Exp. Lung Res. 26:257-271. - PubMed
-
- Berson, D. M., F. A. Dunn, and M. Takao. 2002. Phototransduction by retinal ganglion cells that set the circadian clock. Science 295:1070-1074. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases