

Generation of competent bone marrow-derived antigen presenting cells from the deer mouse (Peromyscus maniculatus)
- PMID: 15458574
- PMCID: PMC524361
- DOI: 10.1186/1471-2172-5-23
Generation of competent bone marrow-derived antigen presenting cells from the deer mouse (Peromyscus maniculatus)
Abstract
Background: Human infections with Sin Nombre virus (SNV) and related New World hantaviruses often lead to hantavirus cardiopulmonary syndrome (HCPS), a sometimes fatal illness. Lungs of patients who die from HCPS exhibit cytokine-producing mononuclear infiltrates and pronounced pulmonary inflammation. Deer mice (Peromyscus maniculatus) are the principal natural hosts of SNV, in which the virus establishes life-long persistence without conspicuous pathology. Little is known about the mechanisms SNV employs to evade the immune response of deer mice, and experimental examination of this question has been difficult because of a lack of methodologies for examining such responses during infection. One such deficiency is our inability to characterize T cell responses because susceptible syngeneic deer mice are not available.
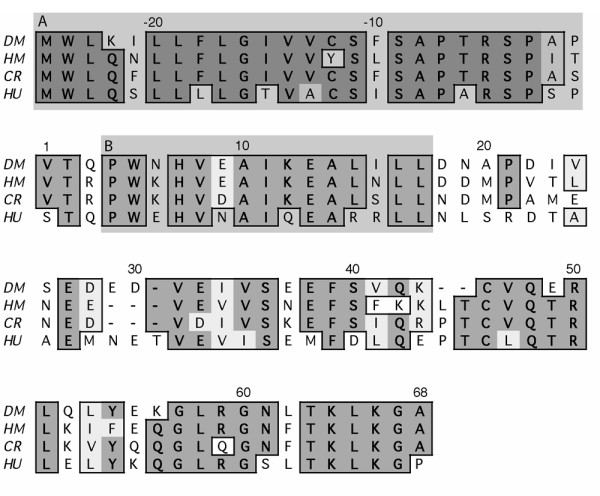
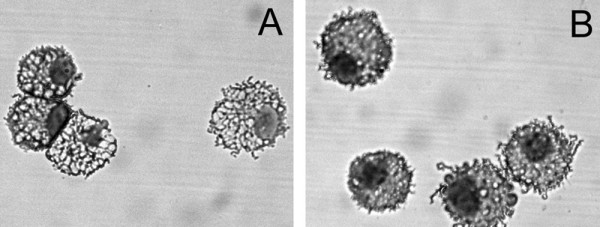
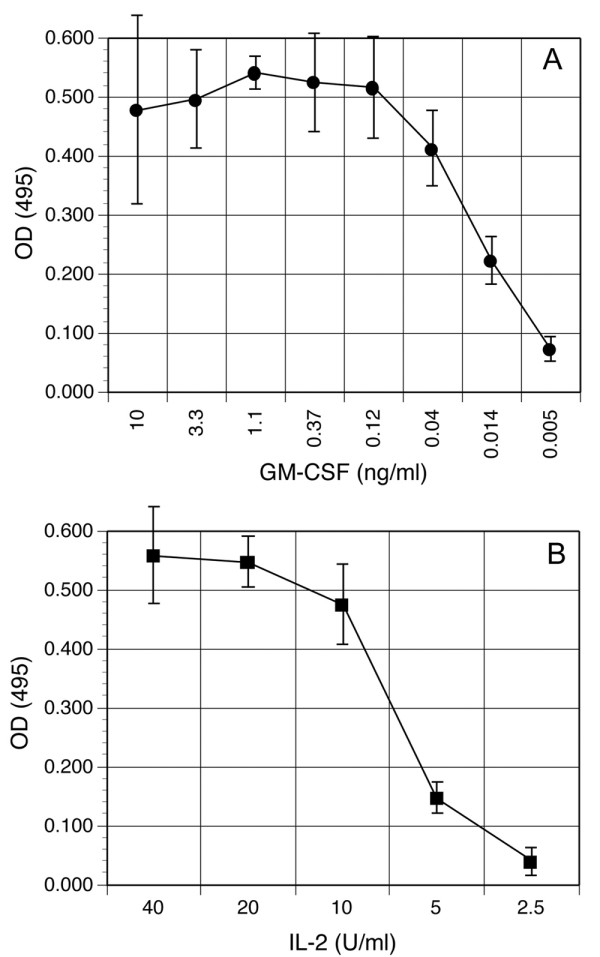
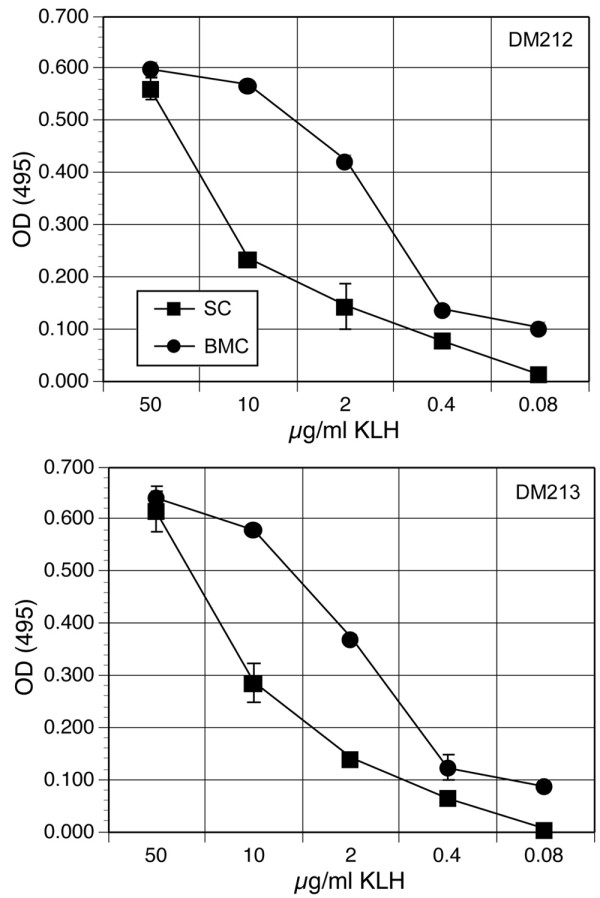
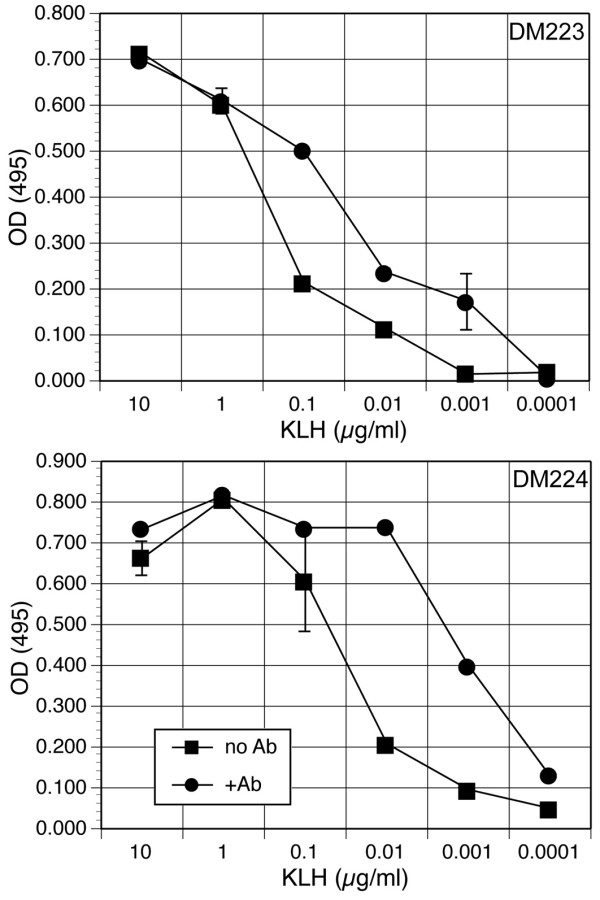
Results: To solve this problem, we have developed an in vitro method of expanding and generating competent antigen presenting cells (APC) from deer mouse bone marrow using commercially-available house mouse (Mus musculus) granulocyte-macrophage colony stimulating factor. These cells are capable of processing and presenting soluble protein to antigen-specific autologous helper T cells in vitro. Inclusion of antigen-specific deer mouse antibody augments T cell stimulation, presumably through Fc receptor-mediated endocytosis.
Conclusions: The use of these APC has allowed us to dramatically expand deer mouse helper T cells in culture and should permit extensive characterization of T cell epitopes. Considering the evolutionary divergence between deer mice and house mice, it is probable that this method will be useful to other investigators using unconventional models of rodent-borne diseases.
Figures
Similar articles
-
Differential lymphocyte and antibody responses in deer mice infected with Sin Nombre hantavirus or Andes hantavirus.J Virol. 2014 Aug;88(15):8319-31. doi: 10.1128/JVI.00004-14. Epub 2014 May 14. J Virol. 2014. PMID: 24829335 Free PMC article.
-
Cloning and characterization of deer mouse (Peromyscus maniculatus) cytokine and chemokine cDNAs.BMC Immunol. 2004 Jan 13;5:1. doi: 10.1186/1471-2172-5-1. BMC Immunol. 2004. PMID: 14720307 Free PMC article.
-
Expression profiling of lymph node cells from deer mice infected with Andes virus.BMC Immunol. 2013 Apr 9;14:18. doi: 10.1186/1471-2172-14-18. BMC Immunol. 2013. PMID: 23570545 Free PMC article.
-
Sin Nombre Virus and the Emergence of Other Hantaviruses: A Review of the Biology, Ecology, and Disease of a Zoonotic Pathogen.Biology (Basel). 2023 Nov 9;12(11):1413. doi: 10.3390/biology12111413. Biology (Basel). 2023. PMID: 37998012 Free PMC article. Review.
-
The Tritope Model for restrictive recognition of antigen by T-cells II. Implications for ontogeny, evolution and physiology.Mol Immunol. 2008 Feb;45(3):632-52. doi: 10.1016/j.molimm.2006.02.033. Epub 2007 Sep 21. Mol Immunol. 2008. PMID: 17889366 Free PMC article. Review.
Cited by
-
Immunology of bats and their viruses: challenges and opportunities.Viruses. 2014 Dec;6(12):4880-901. doi: 10.3390/v6124880. Viruses. 2014. PMID: 25494448 Free PMC article. Review.
-
Use of a Novel Detection Tool to Survey Orthohantaviruses in Wild-Caught Rodent Populations.Viruses. 2022 Mar 25;14(4):682. doi: 10.3390/v14040682. Viruses. 2022. PMID: 35458412 Free PMC article.
-
A human pathogenic hantavirus circulates and is shed in taxonomically diverse rodent reservoirs.PLoS Pathog. 2025 Jan 21;21(1):e1012849. doi: 10.1371/journal.ppat.1012849. eCollection 2025 Jan. PLoS Pathog. 2025. PMID: 39836687 Free PMC article.
-
Hantavirus immunology of rodent reservoirs: current status and future directions.Viruses. 2014 Mar 14;6(3):1317-35. doi: 10.3390/v6031317. Viruses. 2014. PMID: 24638205 Free PMC article. Review.
-
Innate Immunity to Orthohantaviruses: Could Divergent Immune Interactions Explain Host-specific Disease Outcomes?J Mol Biol. 2022 Mar 30;434(6):167230. doi: 10.1016/j.jmb.2021.167230. Epub 2021 Sep 4. J Mol Biol. 2022. PMID: 34487792 Free PMC article. Review.
References
-
- Nichol ST, Spiropoulou CF, Morzunov S, Rollin PE, Ksiazek TG, Feldmann H, Sanchez A, Childs J, Zaki S, Peters CJ. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science. 1993;262:914–917. - PubMed
-
- Hughes JM, Peters CJ, Cohen ML, Mahy BW. Hantavirus pulmonary syndrome: an emerging infectious disease. Science. 1993;262:850–851. - PubMed
-
- Elliott LH, Ksiazek TG, Rollin PE, Spiropoulou CF, Morzunov S, Monroe M, Goldsmith CS, Humphrey CD, Zaki SR, Krebs JW, Maupin GO, Gage KL, Childs J, Nichol S, Peters CJ. Isolation of the causative agent of hantavirus pulmonary syndrome. Am J Trop Med Hyg. 1994;51:102–108. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
