

Review
doi: 10.1038/nrm1488.
Molecular mechanisms of translational control
Affiliations
- PMID: 15459663
- PMCID: PMC7097087
- DOI: 10.1038/nrm1488
Item in Clipboard
Review
Molecular mechanisms of translational control
Nat Rev Mol Cell Biol.
2004 Oct.
Abstract
Translational control is widely used to regulate gene expression. This mode of regulation is especially relevant in situations where transcription is silent or when local control over protein accumulation is required. Although many examples of translational regulation have been described, only a few are beginning to be mechanistically understood. Instead of providing a comprehensive account of the examples that are known at present, we discuss instructive cases that serve as paradigms for different modes of translational control.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
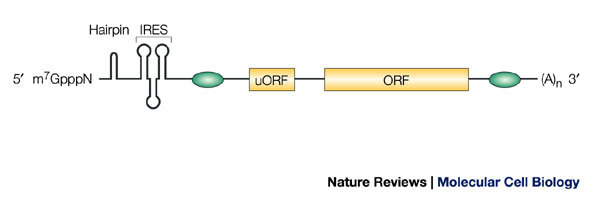
The m7GpppN cap structure at the 5′ end of the mRNA, and the poly(A) tail ((A)n in the figure) at the 3′ end, are canonical motifs that strongly promote translation initiation. Secondary structures, such as hairpins, block translation. Internal ribosome entry sequences (IRESs) mediate cap-independent translation. Upstream open reading frames (uORFs) normally function as negative regulators by reducing translation from the main ORF. Green ovals symbolize binding sites for proteins and/or RNA regulators, which usually inhibit, but occasionally promote, translation.
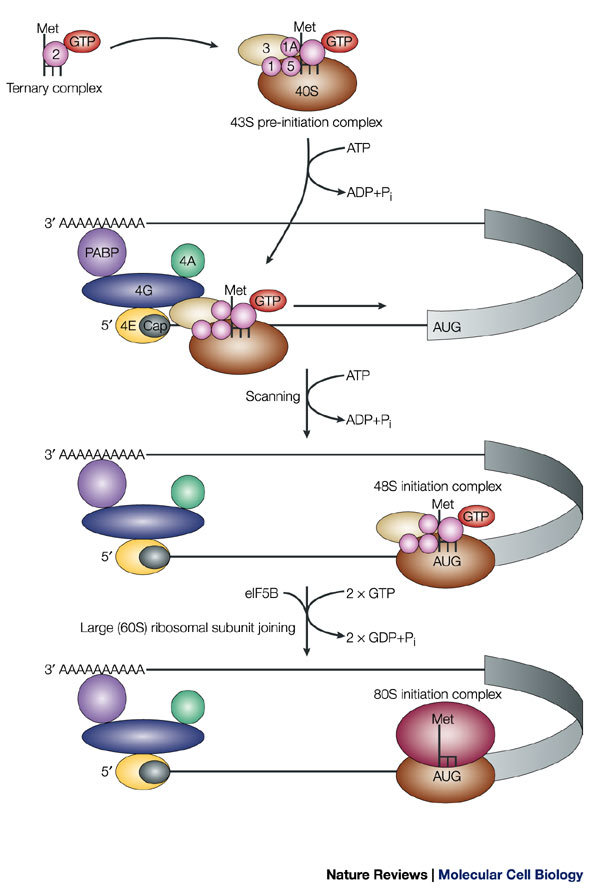
Only the translation-initiation factors that are discussed in the main text are depicted; others have been omitted for simplicity. Eukaryotic initiation factors (eIFs) are depicted as coloured, numbered shapes in the figure. For a complete account of translation-initiation factors, see Refs ,. The methionine-loaded initiator tRNA (L-shaped symbol) binds to GTP-coupled eIF2, to yield the ternary complex. This complex then binds to the small (40S) ribosomal subunit, eIF3 and other initiation factors to form the 43S pre-initiation complex. The pre-initiation complex recognizes the mRNA by the binding of eIF3 to the eIF4G subunit of the cap-binding complex. In addition to eIF4G, the cap-binding complex contains eIF4E, which directly binds to the cap, and eIF4A, an RNA helicase that unwinds secondary structure during the subsequent step of scanning. eIF4G also contacts the poly(A)-binding protein (PABP) and this interaction is thought to circularize the mRNA. The 43S pre-initiation complex scans the mRNA in a 5′→3′ direction until it identifies the initiator codon AUG. Scanning is assisted by the factors eIF1 and eIF1A. Stable binding of the 43S pre-initiation complex to the AUG codon yields the 48S initiation complex. Subsequent joining of the large (60S) ribosomal subunit results in the formation of the 80S initiation complex. Both AUG recognition and joining of the large ribosomal subunit trigger GTP hydrolysis on eIF2 and eIF5B, respectively. Subsequently, the 80S complex is competent to catalyze the formation of the first peptide bond. Pi, inorganic phosphate.
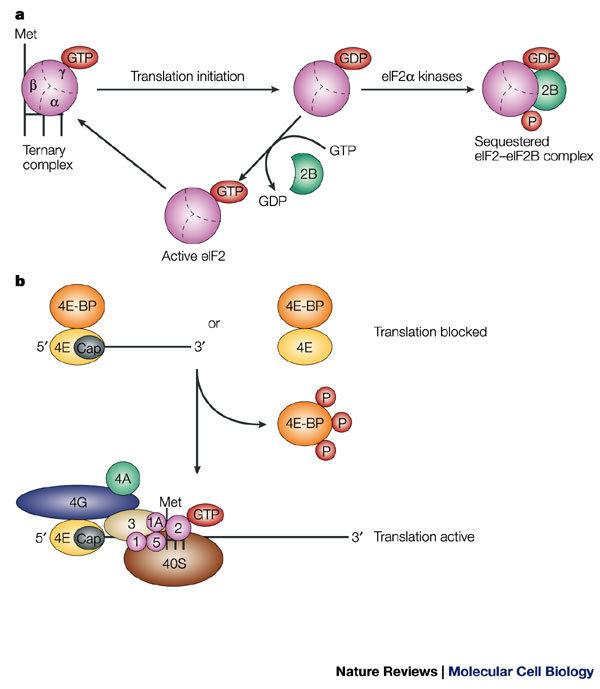
a | GTP hydrolysis and eukaryotic initiation factor (eIF)2 recycling in translation initiation, and the effect of phosphorylation of eIF2α on eIF2 activity. eIF2 consists of three subunits — α, β and γ — and is a component of the ternary complex, which also contains the methionine-loaded initiator tRNA (L-shaped symbol). In an active ternary complex, the eIF2-γ subunit is bound to GTP, and during translation initiation, this GTP molecule is hydrolyzed. GDP–GTP exchange on eIF2 is necessary to re-generate active eIF2 and is catalyzed by eIF2B. Phosphorylation of eIF2 on the α subunit reduces the dissociation rate of eIF2B, thereby sequestering the cellular complement of eIF2B and blocking the GDP–GTP exchange reaction. b | Function of eIF4E-binding proteins (4E-BPs). 4E-BPs bind to eIF4E, thereby preventing its interaction with eIF4G and so inhibiting translation. Phosphorylation of 4E-BP molecules releases the 4E-BPs from eIF4E, which allows their interaction with eIF4G, and thereby allows translation to proceed.
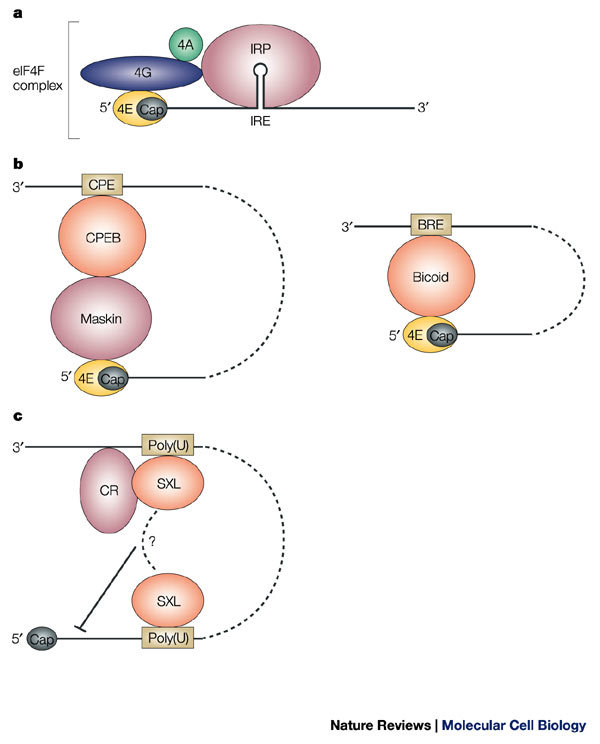
a | Steric blockage. The iron regulatory proteins (IRPs) 1 or 2 bind to the iron-responsive element (IRE) and prevent the recruitment of the 43S pre-initiation complex to the mRNA-bound eukaryotic initiation factor (eIF)4F complex by steric hindrance. b | Interference with the eIF4F complex. The mRNA-specific eIF4E-binding proteins Maskin and Bicoid interact with eIF4E, thereby preventing its interaction with eIF4G. Maskin is targeted to the mRNA through the cytoplasmic-polyadenylation-element-binding protein (CPEB) that recognizes the cytoplasmic polyadenylation element (CPE) that is located at the 3′ untranslated region (UTR), whereas Bicoid directly binds to the mRNA at the Bicoid response element (BRE). c | Cap-independent inhibition. Binding of Sex-lethal (SXL) to uridine-rich sequences (Poly(U) in the figure) at both the 5′ and 3′ UTRs assists the recruitment of a co-repressor complex (CR) to inhibit translation, possibly by interference with ribosome scanning from the cap structure.
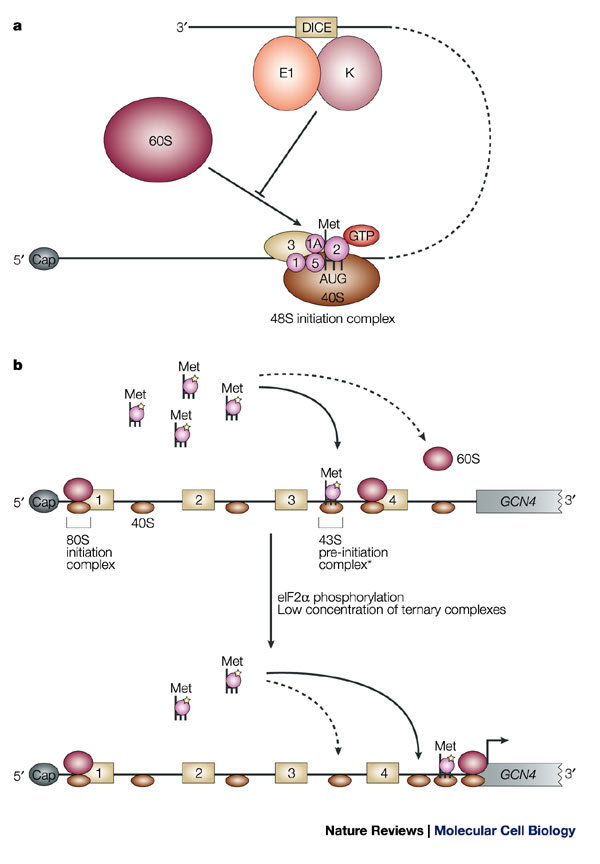
a | Regulation of the association of the 60S ribosomal subunit. Binding of heterogeneous nuclear ribonucleoprotein K (hnRNP K) and hnRNP E1 to the differentiation-control element (DICE) in the 3′ untranslated region (UTR) of LOX mRNA prevents the 60S subunit from joining the 48S initiation complex at the initiator AUG codon. b | Mechanism of regulation of GCN4 mRNA translation. GCN4 mRNA contains four upstream open reading frames (uORF1–4). Under conditions of amino-acid sufficiency (upper panel), reinitiation occurs more frequently after each uORF (continuous arrow), because of an increased probability of recharging the scanning 40S subunits that traverse the regions between the uORFs with active ternary complexes. As a result, reinitiation at the GCN4 ORF becomes infrequent (dashed arrow). Under conditions of amino-acid scarcity, which induces eIF2α phosphorylation, and low levels of ternary complex (lower panel), reinitiation is unlikely to occur at the uORFs. This increases the probability of scanning 40S subunits reaching the region downstream of uORF4 and, subsequently, the GCN4 AUG initiation codon. The asterisk indicates that the exact composition of the 43S pre-initiation complex, in the context of reinitiation, is not known.
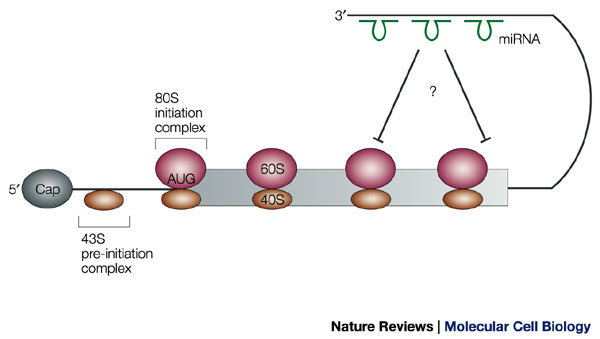
Micro RNAs (miRNAs; shown in green) engage in imperfect base-pairing interactions with the 3′ untranslated region (UTR) and cause translational arrest. At present, evidence indicates that this occurs in polysomal complexes after the initiation of polypeptide synthesis.
References
-
- Wickens M, Goodwin E, Kimble. J, Strickland S, Hentze M. Translational control of gene expression. 2000. p. 295.
-
- Hershey JWB, Merrick WC. Translational control of gene expression. 2000. p. 33.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
