

DNA double strand break repair in human bladder cancer is error prone and involves microhomology-associated end-joining
- PMID: 15466592
- PMCID: PMC521655
- DOI: 10.1093/nar/gkh842
DNA double strand break repair in human bladder cancer is error prone and involves microhomology-associated end-joining
Abstract
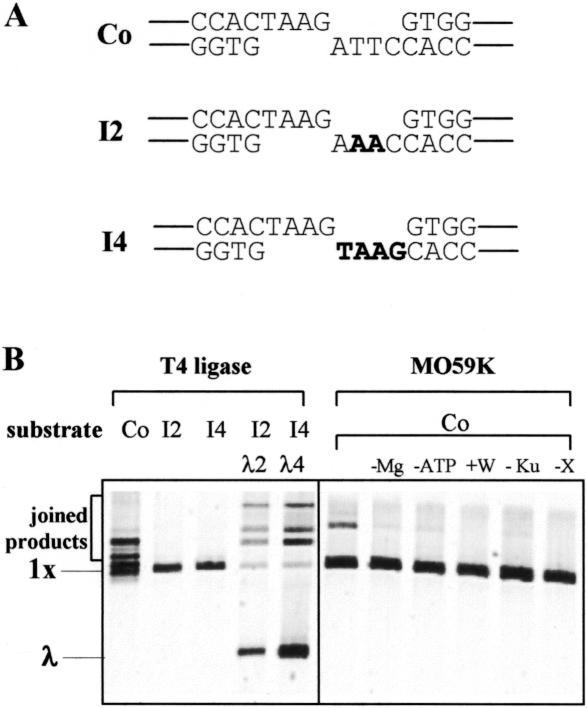
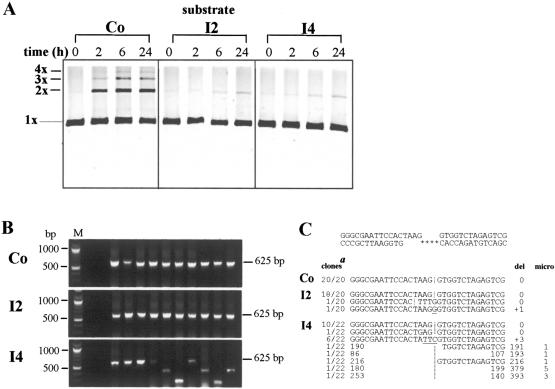
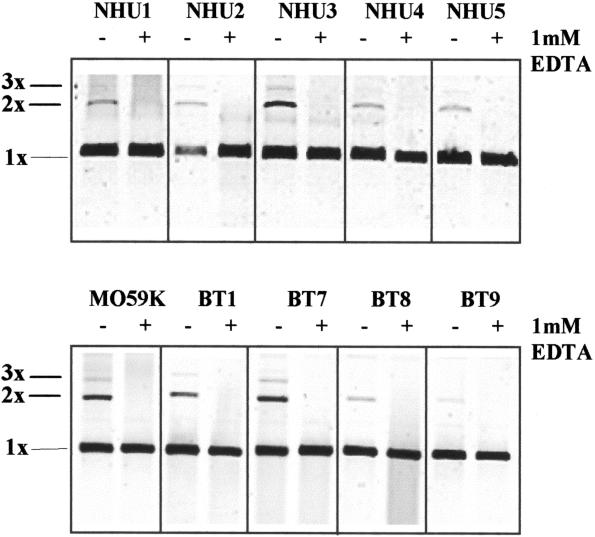
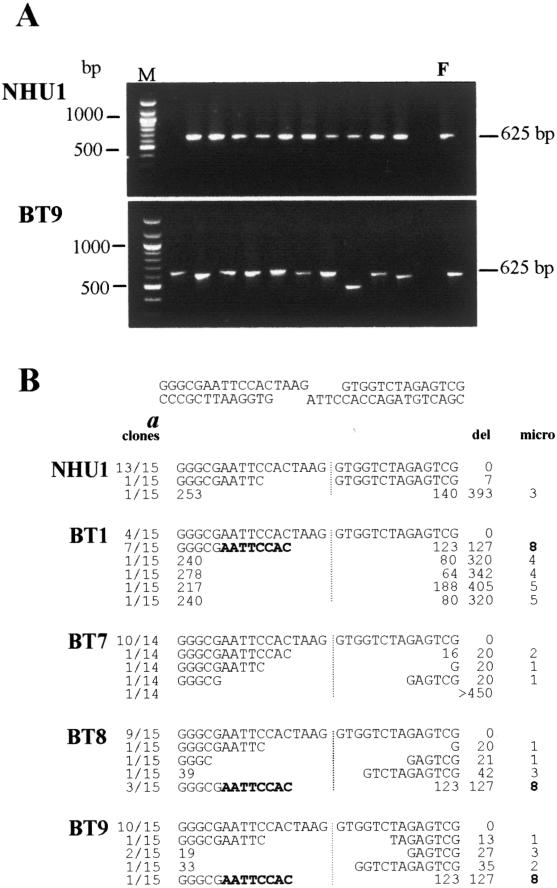
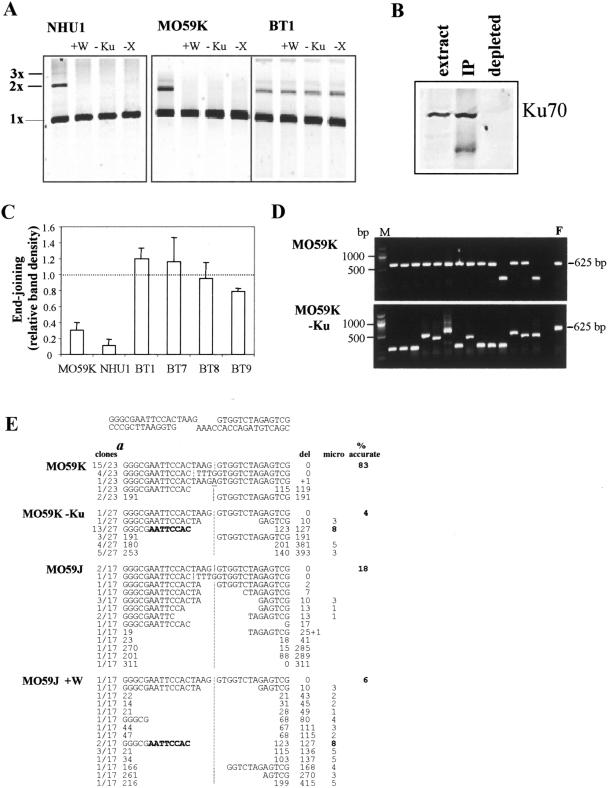
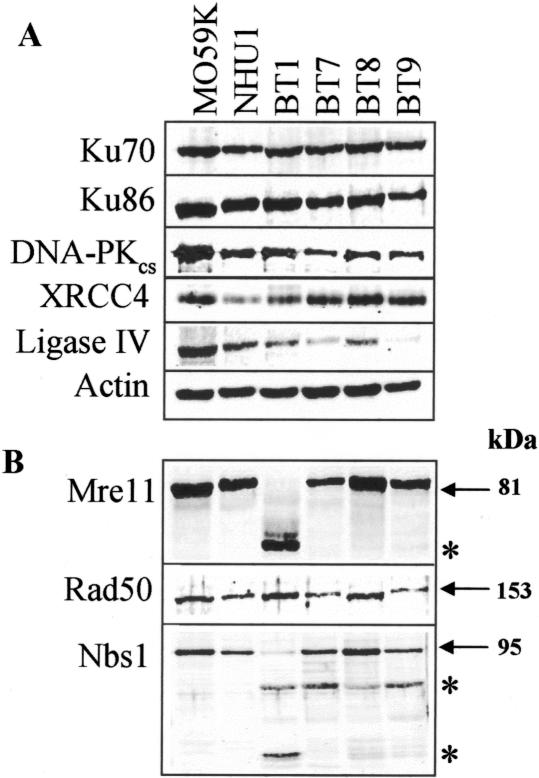
In human cells DNA double strand breaks (DSBs) can be repaired by the non-homologous end-joining (NHEJ) pathway. In a background of NHEJ deficiency, DSBs with mismatched ends can be joined by an error-prone mechanism involving joining between regions of nucleotide microhomology. The majority of joins formed from a DSB with partially incompatible 3' overhangs by cell-free extracts from human glioblastoma (MO59K) and urothelial (NHU) cell lines were accurate and produced by the overlap/fill-in of mismatched termini by NHEJ. However, repair of DSBs by extracts using tissue from four high-grade bladder carcinomas resulted in no accurate join formation. Junctions were formed by the non-random deletion of terminal nucleotides and showed a preference for annealing at a microhomology of 8 nt buried within the DNA substrate; this process was not dependent on functional Ku70, DNA-PK or XRCC4. Junctions were repaired in the same manner in MO59K extracts in which accurate NHEJ was inactivated by inhibition of Ku70 or DNA-PK(cs). These data indicate that bladder tumour extracts are unable to perform accurate NHEJ such that error-prone joining predominates. Therefore, in high-grade tumours mismatched DSBs are repaired by a highly mutagenic, microhomology-mediated, alternative end-joining pathway, a process that may contribute to genomic instability observed in bladder cancer.
Figures

References
-
- Valerie K. and Povirk,L.F. (2003) Regulation and mechanisms of mammalian double strand break repair. Oncogene, 22, 5792–5812. - PubMed
-
- Leiber M.R., Ma,Y., Pannicke,U. and Schwarz,K. (2003) Mechanism and regulation of human non-homologous DNA end-joining. Nature Rev. Mol. Cell. Biol., 4, 712–720. - PubMed
-
- Takata M., Sasaki,M.S., Sonoda,E., Morrison,C., Hashimoto,M., Utsumi,H., Yamaguchi-Iwai,Y., Shinohara,A. and Takeda,S. (1998) Homologous recombination and non-homologous end-joining pathways of DNA double-strand break repair have overlapping roles in the maintenance of chromosomal integrity in vertebrate cells. EMBO J., 17, 5497–5508. - PMC - PubMed
-
- Walker J.R., Corpina,R.A. and Goldberg,J. (2001) Structure of the Ku heterodimer bound to DNA and its implications for double-strand break repair. Nature, 412, 607–614. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
