

doi: 10.1073/pnas.0406410101.
Epub 2004 Oct 4.
The complete genomic sequence of Nocardia farcinica IFM 10152
Affiliations
- PMID: 15466710
- PMCID: PMC522048
- DOI: 10.1073/pnas.0406410101
Item in Clipboard
The complete genomic sequence of Nocardia farcinica IFM 10152
Proc Natl Acad Sci U S A.
.
Abstract
We determined the genomic sequence of Nocardia farcinica IFM 10152, a clinical isolate, and revealed the molecular basis of its versatility. The genome consists of a single circular chromosome of 6,021,225 bp with an average G+C content of 70.8% and two plasmids of 184,027 (pNF1) and 87,093 (pNF2) bp with average G+C contents of 67.2% and 68.4%, respectively. The chromosome encoded 5,674 putative protein-coding sequences, including many candidate genes for virulence and multidrug resistance as well as secondary metabolism. Analyses of paralogous protein families suggest that gene duplications have resulted in a bacterium that can survive not only in soil environments but also in animal tissues, resulting in disease.
Figures
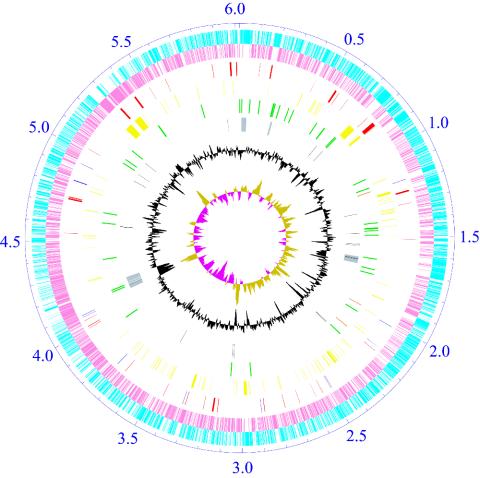
Schematic representation of the N. farcinica chromosome. The ticks show the scale in megabases, with zero representing the location of the dnaA gene. The outer two circles show the predicted protein-coding sequences on the plus (sky blue) and minus (pink) strands; the third shows the putative virulence (red) and drug-resistance (blue) genes; the fourth shows secondary metabolism genes (yellow); the fifth shows rrn operon (orange) and tRNA genes (green); the sixth shows phage-related (gray) and transposase (black) genes; the seventh shows percentage G+C in relation to mean G+C for the chromosome; and the eighth shows GC-skew (khaki, >0; purple, <0).
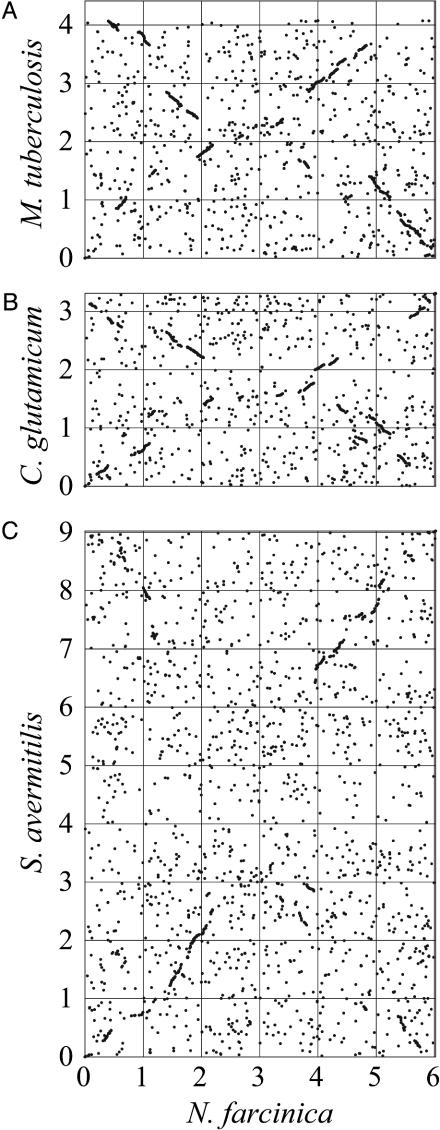
Ortholog plots of N. farcinica vs. M. tuberculosis (A), C. glutamicum (B), or S. avermitilis (C). Orthologs were identified as reciprocal best hits by using the blastp program. For each genome, the dnaA gene is located at position zero.
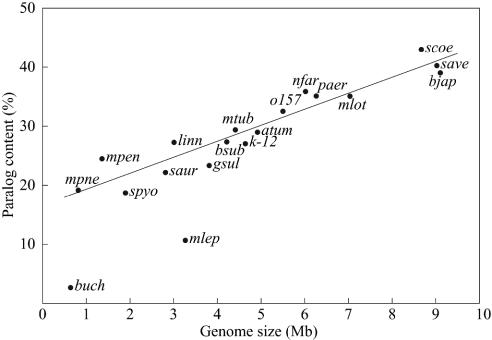
Correlation between paralog content and genome size in various bacteria: atum, Agrobacterium tumefaciens; bjap, Bradyrhizobium japonicum; bsub, Bacillus subtilis; buch, Buchnera sp.; o157, Escherichia coli O-157; k-12, E. coli K-12; gsul, Geobacter sulfurreducens; linn, Listeria innocua; mlot, Mesorhizobium loti; mtub, M. tuberculosis; mlep, Mycobacterium leprae; mpen, Mycoplasma penetrans; mpne, Mycoplasma pneumoniae; nfar, N. farcinica; paer, Pseudomonas aeruginosa; saur, Staphylococcus aureus; spyo, Streptococcus pyogenes; save, S. avermitilis; and scoe, S. coelicolor A3 (2). A linear regression line, which was estimated without using buch and mlep data, is also indicated. Extremely low paralogies in buch and mlep are due to massive decay of genes in these genomes.
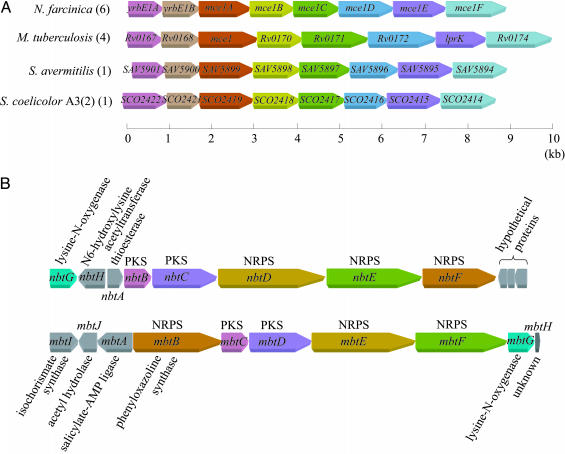
Conservation of gene organization in actinomycetes. (A) Conservation of mce operons. Orthologous genes are denoted by the same color. Numbers of operons found in each genome are indicated in parentheses. (B) Comparison of gene organization between the putative siderophore (Upper) and the mycobactin (Lower) biosynthesis gene clusters. Orthologous and nonorthologous genes are denoted by the same color or in gray, respectively. Both clusters contain three nonribosomal peptide synthetases (NRPSs) and two polyketide synthases (PKSs).
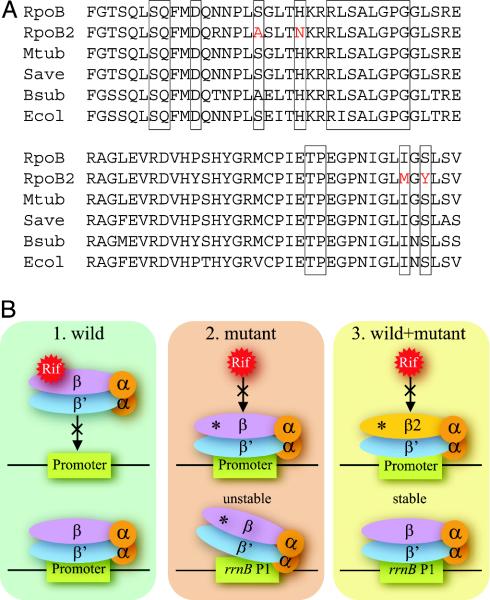
(A) Comparison of the rifampin regions of RNAP β subunits among N. farcinica (RpoB and RpoB2), M. tuberculosis (Mtub), S. avermitilis (Save), B. subtilis (Bsub), and E. coli (Ecol). Boxed regions indicate known positions where mutations lead to rifampin resistance in various bacteria. Substitutions found in the RpoB2 are red-colored. (B) Possible advantage of two types of rpoB genes being present in one genome. 1, Rifampin binds to wild-type RNAP and inhibits its function. 2, Certain mutations (asterisks) on the rpoB gene diminish the binding of rifampin to RNAP, making bacteria resistant in the presence of rifampin. However, rpoB mutants destabilize initiation complexes between RNAP and stringently controlled promoters, for example rrnB P1, even in the absence of rifampin. 3, If both types of rpoB genes are present in one genome, such side effects disappear in the absence of rifampin by the wild-type molecule, and bacteria can survive in the presence of rifampin by the mutant molecule.
Similar articles
-
Analysis of the complete genome sequence of Nocardia seriolae UTF1, the causative agent of fish nocardiosis: The first reference genome sequence of the fish pathogenic Nocardia species.PLoS One. 2017 Mar 3;12(3):e0173198. doi: 10.1371/journal.pone.0173198. eCollection 2017. PLoS One. 2017. PMID: 28257489 Free PMC article.
-
Contribution of rpoB2 RNA polymerase beta subunit gene to rifampin resistance in Nocardia species.Antimicrob Agents Chemother. 2006 Apr;50(4):1342-6. doi: 10.1128/AAC.50.4.1342-1346.2006. Antimicrob Agents Chemother. 2006. PMID: 16569850 Free PMC article.
-
Complete genome sequence analysis of Nocardia brasiliensis HUJEG-1 reveals a saprobic lifestyle and the genes needed for human pathogenesis.PLoS One. 2013 Jun 3;8(6):e65425. doi: 10.1371/journal.pone.0065425. Print 2013. PLoS One. 2013. PMID: 23755230 Free PMC article.
-
[Recent progress in taxonomic studies on pathogenic nocardia and usefulness of the bacteria for the studies on secondary metabolites and antibiotic resistant mechanisms].Nihon Ishinkin Gakkai Zasshi. 2010;51(4):179-92. doi: 10.3314/jjmm.51.179. Nihon Ishinkin Gakkai Zasshi. 2010. PMID: 21060211 Review. Japanese.
-
Borrelia genomes in the year 2000.J Mol Microbiol Biotechnol. 2000 Oct;2(4):401-10. J Mol Microbiol Biotechnol. 2000. PMID: 11075912 Review.
Cited by
-
The Nocardia cyriacigeorgica GUH-2 genome shows ongoing adaptation of an environmental Actinobacteria to a pathogen's lifestyle.BMC Genomics. 2013 Apr 27;14:286. doi: 10.1186/1471-2164-14-286. BMC Genomics. 2013. PMID: 23622346 Free PMC article.
-
Genome sequence of the human- and animal-pathogenic strain Nocardia cyriacigeorgica GUH-2.J Bacteriol. 2012 Apr;194(8):2098-9. doi: 10.1128/JB.00161-12. J Bacteriol. 2012. PMID: 22461543 Free PMC article.
-
Small but sufficient: the Rhodococcus phage RRH1 has the smallest known Siphoviridae genome at 14.2 kilobases.J Virol. 2012 Jan;86(1):358-63. doi: 10.1128/JVI.05460-11. Epub 2011 Oct 19. J Virol. 2012. PMID: 22013058 Free PMC article.
-
Cloning and characterization of alpha-methylacyl coenzyme A racemase from Gordonia polyisoprenivorans VH2.Appl Environ Microbiol. 2008 Nov;74(22):7085-9. doi: 10.1128/AEM.01491-08. Epub 2008 Sep 26. Appl Environ Microbiol. 2008. PMID: 18820059 Free PMC article.
-
Genome analysis reveals three genomospecies in Mycobacterium abscessus.BMC Genomics. 2014 May 12;15(1):359. doi: 10.1186/1471-2164-15-359. BMC Genomics. 2014. PMID: 24886480 Free PMC article.
References
-
- Brown, J. M., McNeil, M. M. & Desmond, E. P. (1999) in Manual of Clinical Microbiology, eds. Murray, P. R., Baron, E. J., Pfaller, M. A., Tenover, F. C. & Yolken, R. H. (Am. Soc. Microbiol., Washington, DC), pp. 370-398.
-
- Kageyama, A., Yazawa, K., Ishikawa, J., Hotta, K., Nishimura, K. & Mikami, Y. (2004) Eur. J. Epidemiol. 19, 383-389. - PubMed
-
- Shigemori, H., Komaki, H., Yazawa, K., Mikami, Y., Nemoto, A., Tanaka, Y., Sasaki, Y., Ishida, T. & Kobayashi, J. (1998) J. Org. Chem. 63, 6900-6904. - PubMed
-
- Tanaka, Y., Komaki, H., Yazawa, K., Mikami, Y., Nemoto, A., Tojyo, T., Kadowaki, K., Shigemori, H. & Kobayashi, J. (1997) J. Antibiot. 50, 1036-1041. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
