

Vasohibin as an endothelium-derived negative feedback regulator of angiogenesis
- PMID: 15467828
- PMCID: PMC518662
- DOI: 10.1172/JCI21152
Vasohibin as an endothelium-derived negative feedback regulator of angiogenesis
Abstract
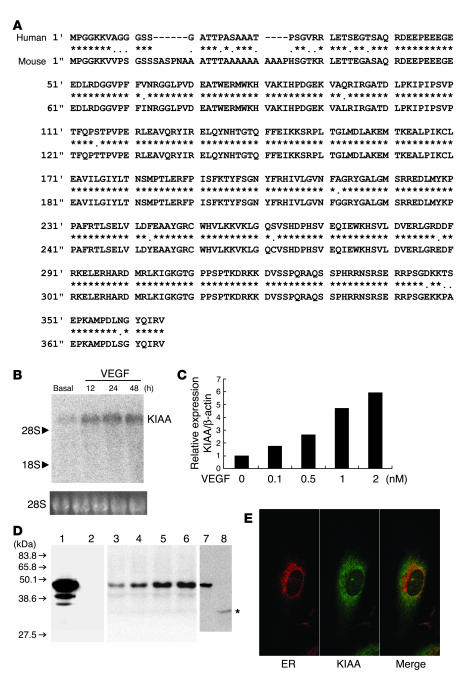
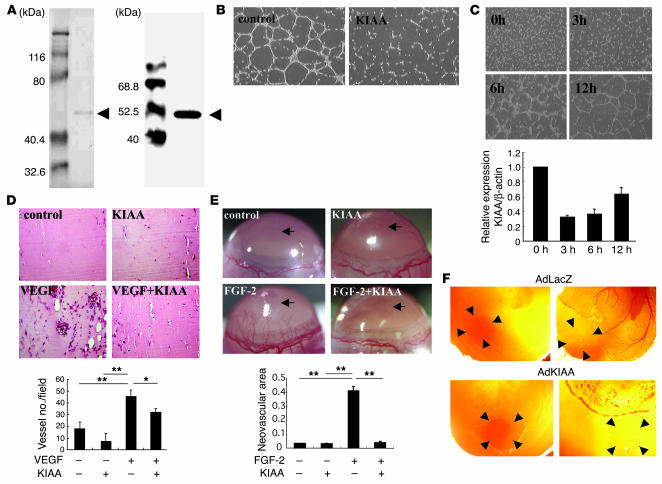
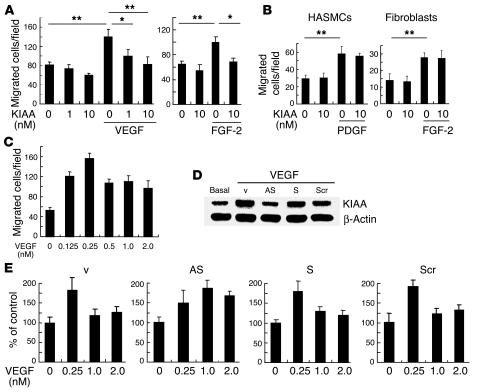
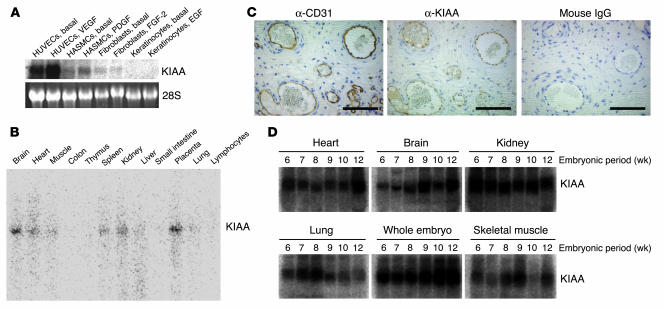
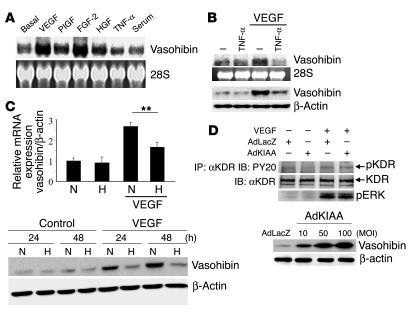
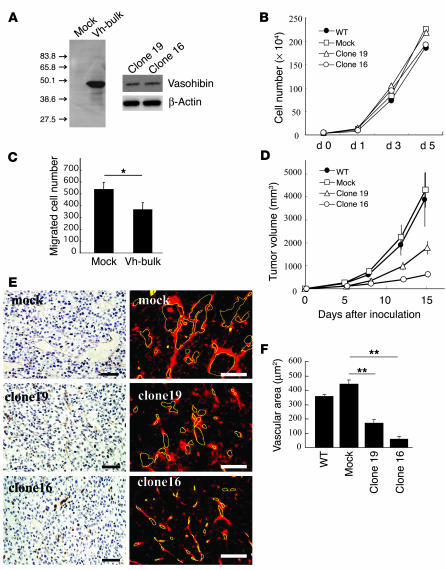
Negative feedback is a crucial physiological regulatory mechanism, but no such regulator of angiogenesis has been established. Here we report a novel angiogenesis inhibitor that is induced in endothelial cells (ECs) by angiogenic factors and inhibits angiogenesis in an autocrine manner. We have performed cDNA microarray analysis to survey VEGF-inducible genes in human ECs. We characterized one such gene, KIAA1036, whose function had been uncharacterized. The recombinant protein inhibited migration, proliferation, and network formation by ECs as well as angiogenesis in vivo. This inhibitory effect was selective to ECs, as the protein did not affect the migration of smooth muscle cells or fibroblasts. Specific elimination of the expression of KIAA1036 in ECs restored their responsiveness to a higher concentration of VEGF. The expression of KIAA1036 was selective to ECs, and hypoxia or TNF-alpha abrogated its inducible expression. As this molecule is preferentially expressed in ECs, we designated it "vasohibin." Transfection of Lewis lung carcinoma cells with the vasohibin gene did not affect the proliferation of cancer cells in vitro, but did inhibit tumor growth and tumor angiogenesis in vivo. We propose vasohibin to be an endothelium-derived negative feedback regulator of angiogenesis.
Figures
Comment in
-
Vasohibin: the feedback on a new inhibitor of angiogenesis.J Clin Invest. 2004 Oct;114(7):884-6. doi: 10.1172/JCI23153. J Clin Invest. 2004. PMID: 15467823 Free PMC article.
References
-
- Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat. Med. 1995;1:27–31. - PubMed
-
- Ribatti D, Vacca A, Presta M. The discovery of angiogenic factors: a historical review. Gen. Pharmacol. 2000;35:227–231. - PubMed
-
- Ferrara N. VEGF and the quest for tumour angiogenesis factors. Nat. Rev. Cancer. 2002;2:795–803. - PubMed
-
- Dameron KM, Volpert OV, Tainsky MA, Bouck NP. Control of angiogenesis in fibroblasts by p53 regulation of thrombospondin-1. Science. 1994;265:1582–1584. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
