

CEACAM1 modulates epidermal growth factor receptor--mediated cell proliferation
- PMID: 15467833
- PMCID: PMC518664
- DOI: 10.1172/JCI21786
CEACAM1 modulates epidermal growth factor receptor--mediated cell proliferation
Abstract
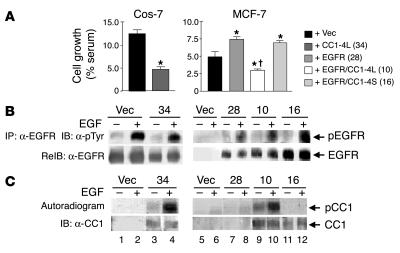
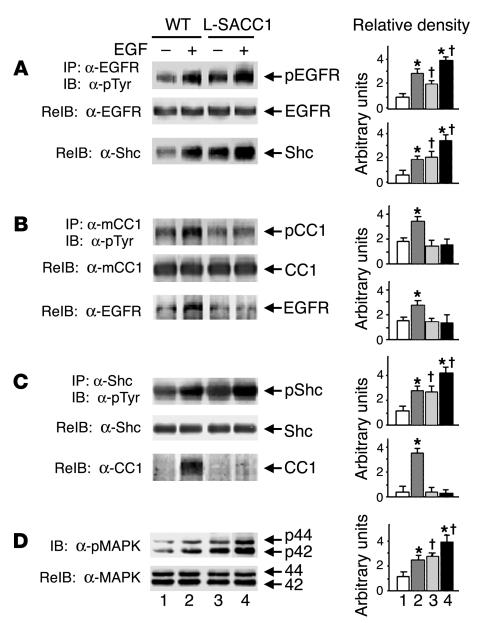
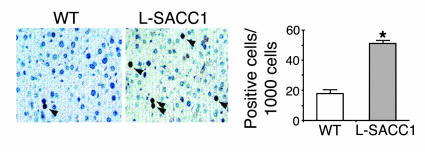
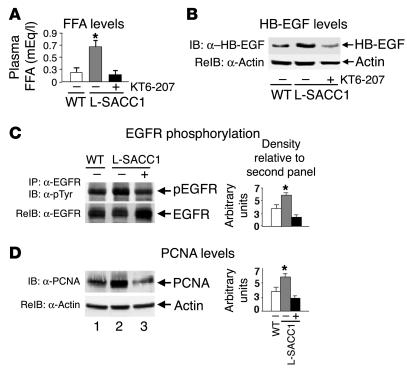
Phosphorylation of the cell adhesion protein CEACAM1 increases insulin sensitivity and decreases insulin-dependent mitogenesis in vivo. Here we show that CEACAM1 is a substrate of the EGFR and that upon being phosphorylated, CEACAM1 reduces EGFR-mediated growth of transfected Cos-7 and MCF-7 cells in response to EGF. Using transgenic mice overexpressing a phosphorylation-defective CEACAM1 mutant in liver (L-SACC1), we show that the effect of CEACAM1 on EGF-dependent cell proliferation is mediated by its ability to bind to and sequester Shc, thus uncoupling EGFR signaling from the ras/MAPK pathway. In L-SACC1 mice, we also show that impaired CEACAM1 phosphorylation leads to ligand-independent increase of EGFR-mediated cell proliferation. This appears to be secondary to visceral obesity and the metabolic syndrome, with increased levels of output of free fatty acids and heparin-binding EGF-like growth factor from the adipose tissue of the mice. Thus, L-SACC1 mice provide a model for the mechanistic link between increased cell proliferation in states of impaired metabolism and visceral obesity.
Figures
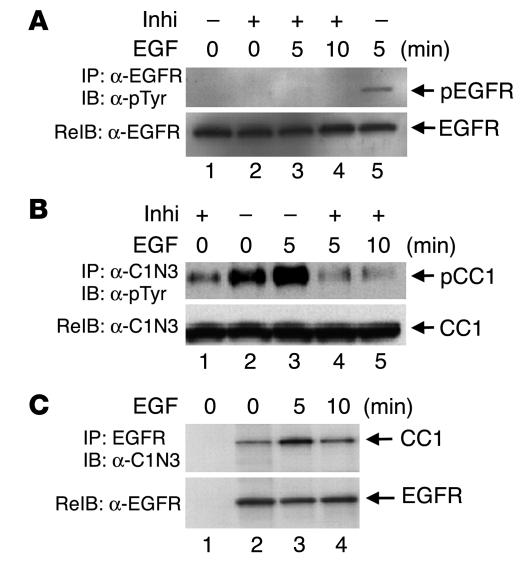
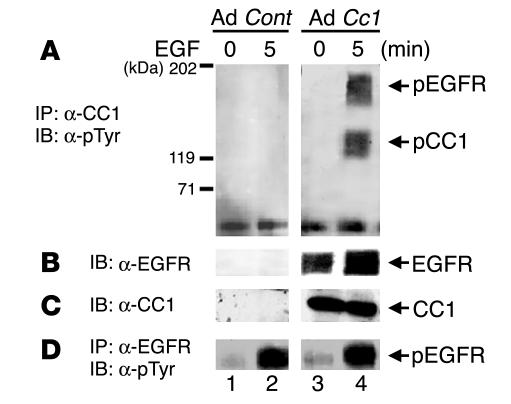
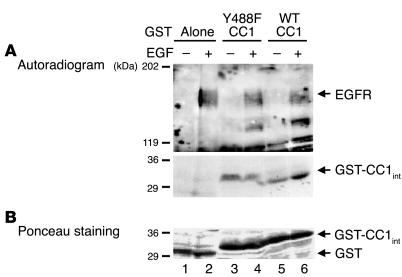
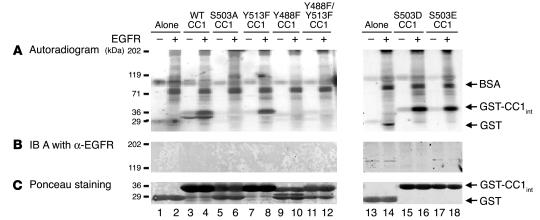
References
-
- Crews CM, Erikson RL. Extracellular signals and reversible protein phosphorylation: what to Mek of it all. Cell. 1993;74:215–217. - PubMed
-
- Cantley LC. The phosphoinositide 3-kinase pathway [review] Science. 2002;296:1655–1657. - PubMed
-
- Wagener C, Ergun S. Angiogenic properties of the carcinoembryonic antigen-related cell adhesion molecule. Exp. Cell Res. 2000;261:19–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
