

Activation of IKKalpha target genes depends on recognition of specific kappaB binding sites by RelB:p52 dimers
- PMID: 15470505
- PMCID: PMC524385
- DOI: 10.1038/sj.emboj.7600391
Activation of IKKalpha target genes depends on recognition of specific kappaB binding sites by RelB:p52 dimers
Abstract
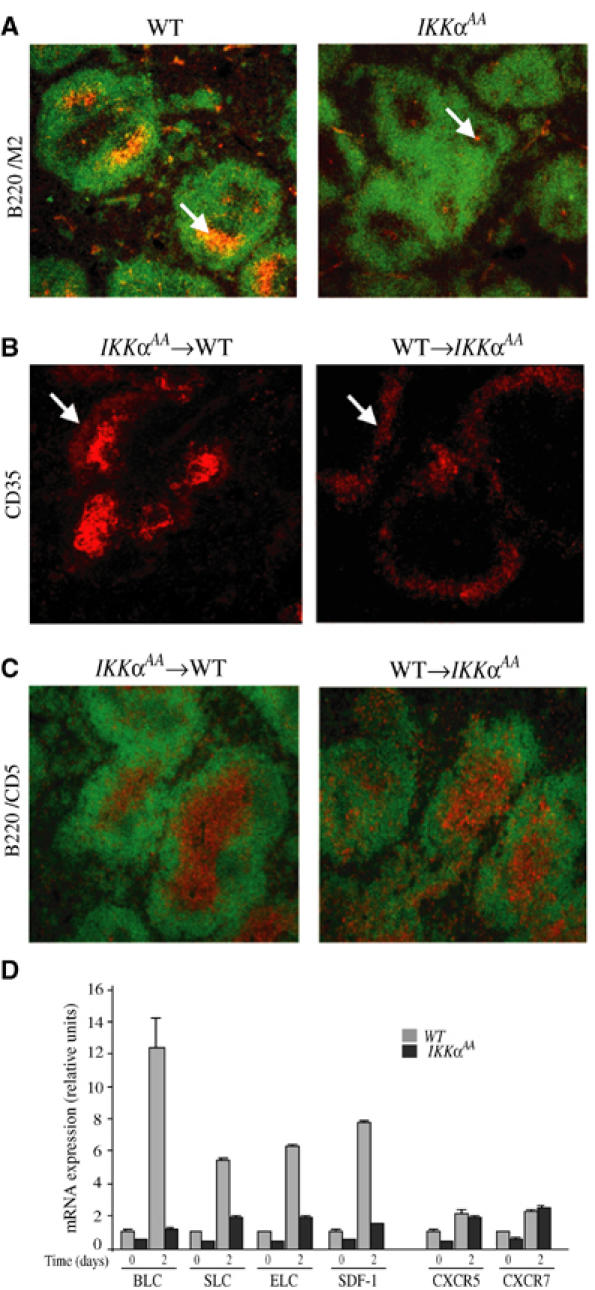
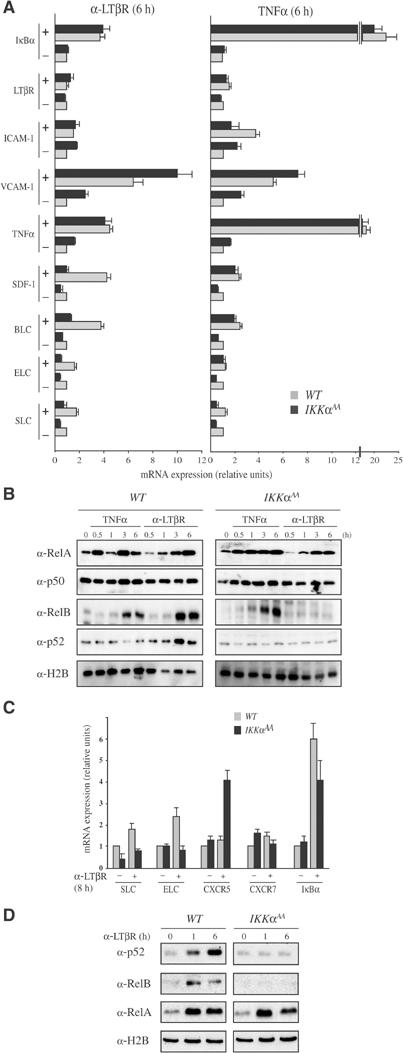
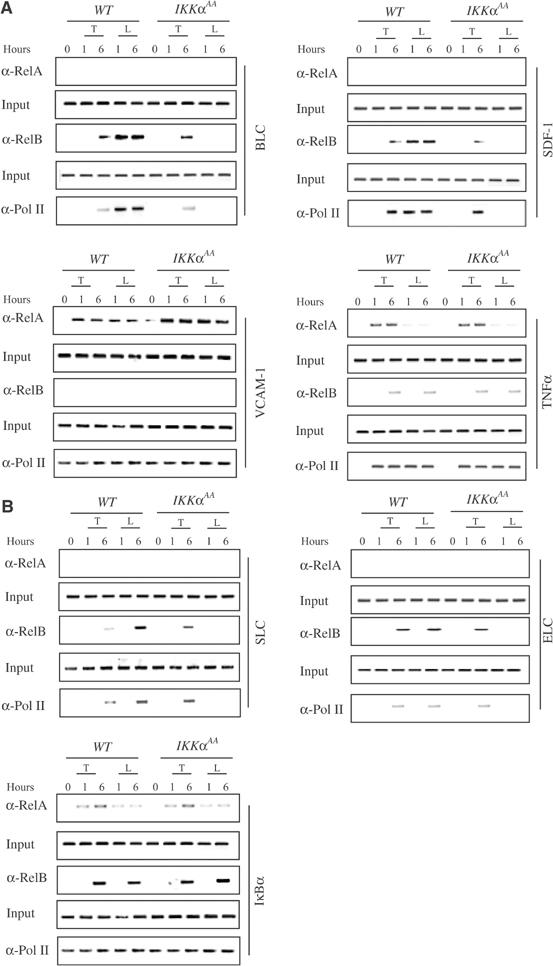
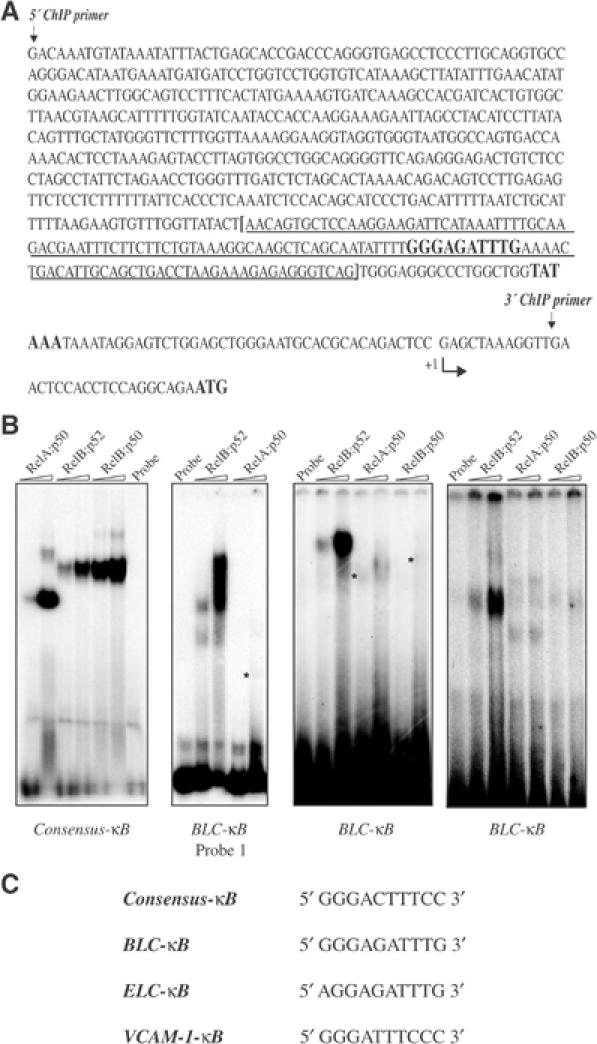
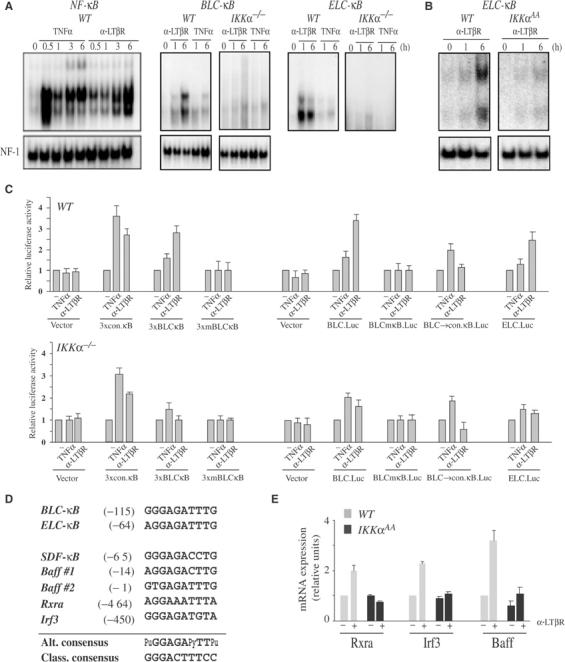
IkappaB Kinase (IKK)alpha is required for activation of an alternative NF-kappaB signaling pathway based on processing of the NF-kappaB2/p100 precursor protein, which associates with RelB in the cytoplasm. This pathway, which activates RelB:p52 dimers, is required for induction of several chemokine genes needed for organization of secondary lymphoid organs. We investigated the basis for the IKKalpha dependence of the induction of these genes in response to engagement of the lymphotoxin beta receptor (LTbetaR). Using chromatin immunoprecipitation, we found that the promoters of organogenic chemokine genes are recognized by RelB:p52 dimers and not by RelA:p50 dimers, the ubiquitous target for the classical NF-kappaB signaling pathway. We identified in the IKKalpha-dependent promoters a novel type of NF-kappaB-binding site that is preferentially recognized by RelB:p52 dimers. This site links induction of organogenic chemokines and other important regulatory molecules to activation of the alternative pathway.
Figures
References
-
- Alcamo E, Mizgerd JP, Horwitz BH, Bronson R, Beg AA, Scott M, Doerschuk CM, Hynes RO, Baltimore D (2001) Targeted mutation of TNF receptor I rescues the RelA-deficient mouse and reveals a critical role for NF-κB in leukocyte recruitment. J Immunol 167: 1592–1600 - PubMed
-
- Anest V, Hanson JL, Cogswell PC, Steinbrecher KA, Strahl BD, Baldwin AS (2003) A nucleosomal function for IκB kinase-α in NF-κB-dependent gene expression. Nature 423: 659–663 - PubMed
-
- Ansel KM, Cyster JG (2001) Chemokines in lymphopoiesis and lymphoid organ development. Curr Opin Immunol 13: 172–179 - PubMed
-
- Ansel KM, Ngo VN, Hyman PL, Luther SA, Forster R, Sedgwick JD, Browning JL, Lipp M, Cyster JG (2000) A chemokine-driven positive feedback loop organizes lymphoid follicles. Nature 406: 309–314 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
