

Iron-induced oligomerization of yeast frataxin homologue Yfh1 is dispensable in vivo
- PMID: 15472712
- PMCID: PMC1299167
- DOI: 10.1038/sj.embor.7400272
Iron-induced oligomerization of yeast frataxin homologue Yfh1 is dispensable in vivo
Abstract
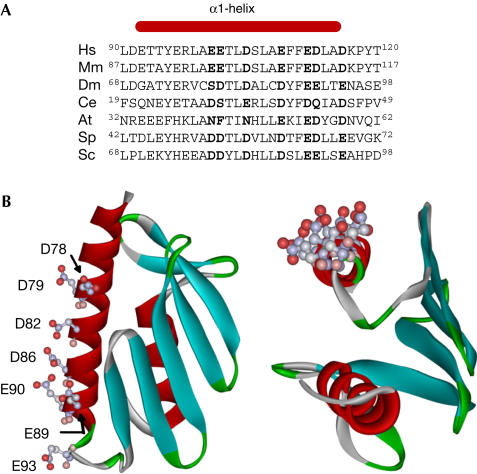
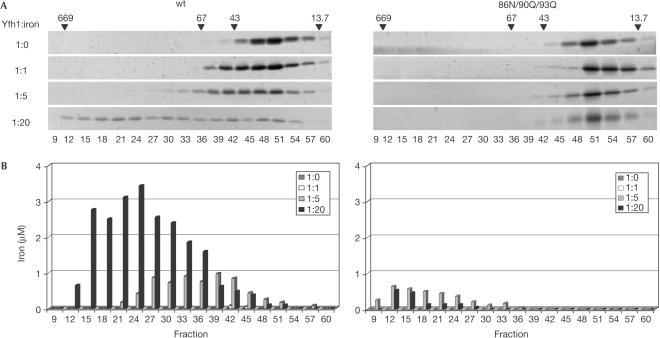
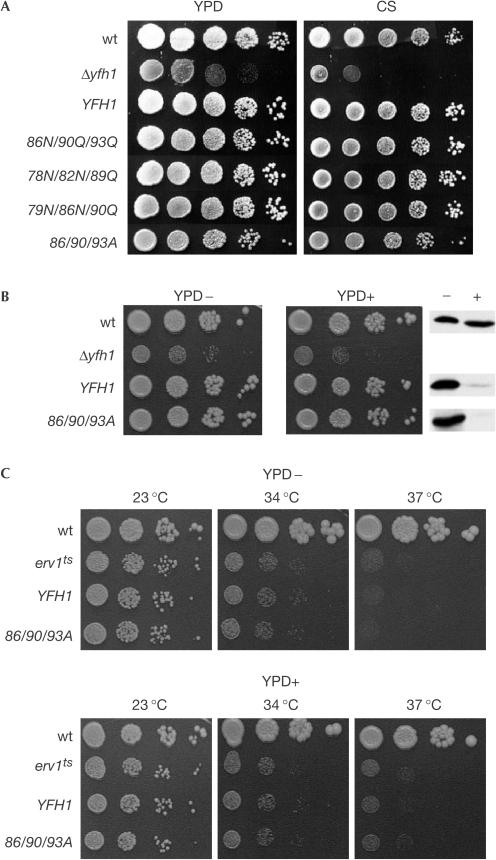
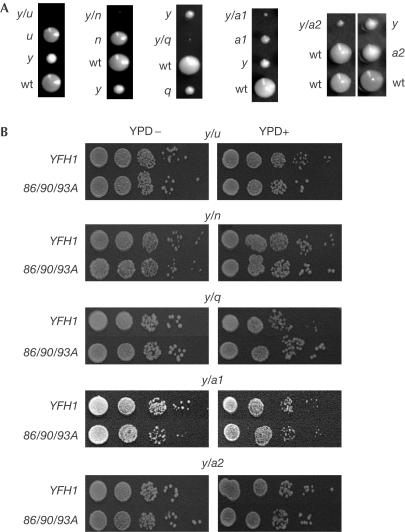
The neurodegenerative disease Friedreich's ataxia is caused by reduced levels of frataxin, a mitochondrial matrix protein. The in vivo role of frataxin is under debate. Frataxin, as well as its yeast homologue Yfh1, binds multiple iron atoms as an oligomer and has been proposed to function as a crucial iron-storage protein. We identified a mutant Yfh1 defective in iron-induced oligomerization. This mutant protein was able to replace functionally wild-type Yfh1, even when expressed at low levels, when mitochondrial iron levels were high and in mutant strains having deletions of genes that had synthetic growth defects with a YFH1 deletion. The ability of an oligomerization-deficient Yfh1 to function in vivo suggests that oligomerization, and thus oligomerization-induced iron storage, is not a critical function of Yfh1. Rather, the capacity of this oligomerization-deficient mutant to interact with the Isu protein suggests a more direct role of Yfh1 in iron-sulphur cluster biogenesis.
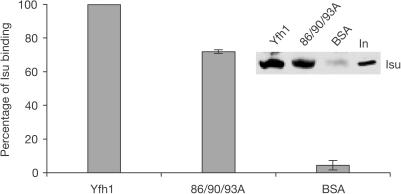
Figures
References
-
- Adinolfi S, Trifuoggi M, Politou AS, Martin S, Pastore A (2002) A structural approach to understanding the iron-binding properties of phylogenetically different frataxins. Hum Mol Genet 11: 1865–1877 - PubMed
-
- Babcock M, de Silva D, Oaks R, Davis-Kaplan S, Jiralerspong S, Montermini L, Pandolfo M, Kaplan J (1997) Regulation of mitochondrial iron accumulation by Yfh1p, a putative homolog of frataxin. Science 276: 1709–1712 - PubMed
-
- Cavadini P, O'Neill HA, Benada O, Isaya G (2002) Assembly and iron-binding properties of human frataxin, the protein deficient in Friedreich ataxia. Hum Mol Genet 11: 217–227 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
