

Hepatic farnesyl diphosphate synthase expression is suppressed by polyunsaturated fatty acids
- PMID: 15473864
- PMCID: PMC1134755
- DOI: 10.1042/BJ20040933
Hepatic farnesyl diphosphate synthase expression is suppressed by polyunsaturated fatty acids
Abstract
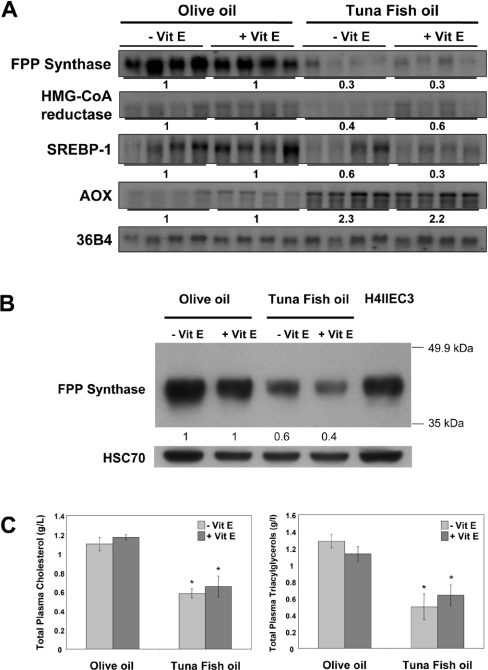
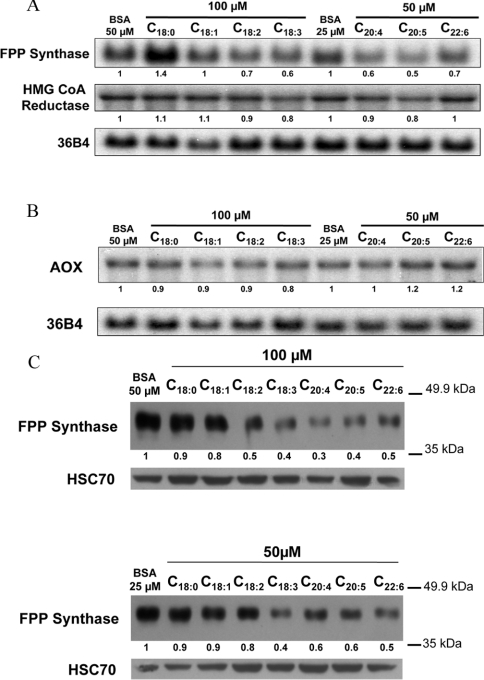
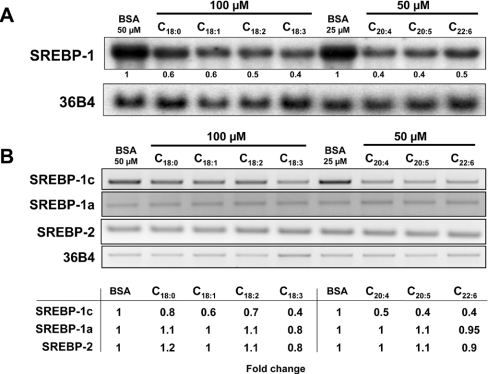
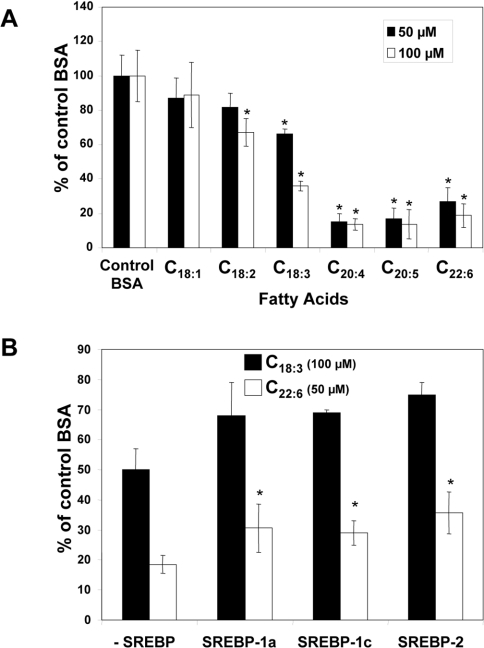
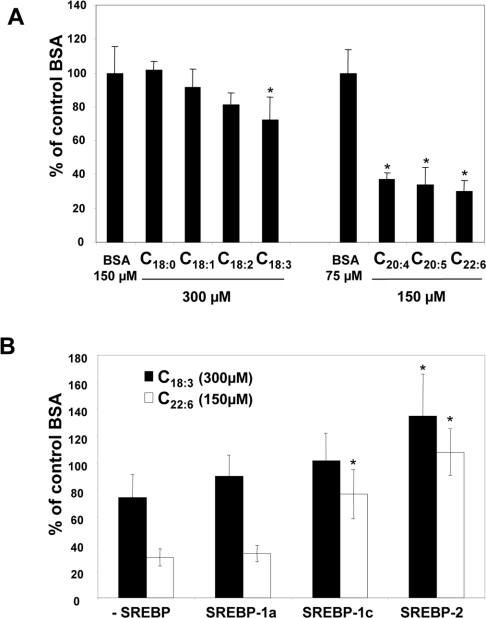
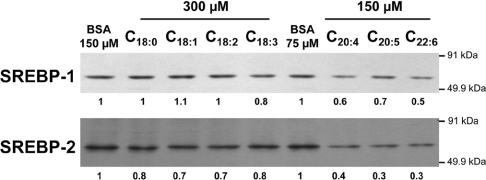
Dietary vegetable oils and fish oils rich in PUFA (polyunsaturated fatty acids) exert hypocholesterolaemic and hypotriglyceridaemic effects in rodents. The plasma cholesterol-lowering properties of PUFA are due partly to a diminution of cholesterol synthesis and of the activity of the rate-limiting enzyme HMG-CoA reductase (3-hydroxy-3-methylglutaryl-CoA reductase). To better understand the mechanisms involved, we examined how tuna fish oil and individual n-3 and n-6 PUFA affect the expression of hepatic FPP synthase (farnesyl diphosphate synthase), a SREBP (sterol regulatory element-binding protein) target enzyme that is subject to negative-feedback regulation by sterols, in co-ordination with HMG-CoA reductase. Feeding mice on a tuna fish oil diet for 2 weeks decreased serum cholesterol and triacylglycerol levels, by 50% and 60% respectively. Hepatic levels of FPP synthase and HMG-CoA reductase mRNAs were also decreased, by 70% and 40% respectively. Individual n-3 and n-6 PUFA lowered FPP synthase and HMG-CoA reductase mRNA levels in H4IIEC3 rat hepatoma cells to a greater extent than did stearate and oleate, with the largest inhibitory effects occurring with arachidonate, EPA (eicosapentaenoic acid) and DHA (docosahexaenoic acid). We observed a similar inhibitory effect on protein levels of FPP synthase. The suppressive effect of PUFA on the FPP synthase mRNA level was not due to a decrease in mRNA stability, but to transcription inhibition. Moreover, a lower nuclear availability of both SREBP-1 and SREBP-2 mature forms was observed in HepG2 human hepatoblastoma cells treated with arachidonate, EPA or DHA. Taken together, these data suggest that PUFA can down-regulate hepatic cholesterol synthesis through inhibition of HMG-CoA reductase and FPP synthase, at least in part through impairment of the SREBP pathway.
Figures

References
-
- Ross R. The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature (London) 1993;362:801–809. - PubMed
-
- Angerer P., von Schacky C. n-3 polyunsaturated fatty acids and the cardiovascular system. Curr. Opin. Lipidol. 2000;1:57–63. - PubMed
-
- Harris W. S. Fish oils and plasma lipid and lipoprotein metabolism in humans: a critical review. J. Lipid Res. 1989;6:785–807. - PubMed
-
- Nestel P. J. Effects of n-3 fatty acids on lipid metabolism. Annu. Rev. Nutr. 1990;10:149–167. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials