

Evolutionary transition toward defective RNAs that are infectious by complementation
- PMID: 15479809
- PMCID: PMC523252
- DOI: 10.1128/JVI.78.21.11678-11685.2004
Evolutionary transition toward defective RNAs that are infectious by complementation
Abstract
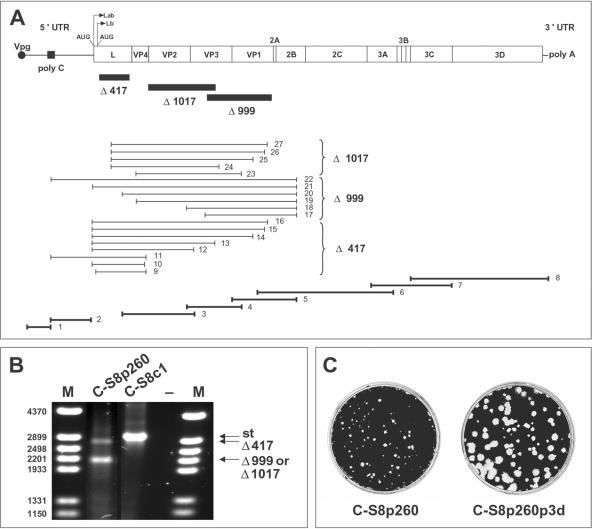
Passage of foot-and-mouth disease virus (FMDV) in cell culture resulted in the generation of defective RNAs that were infectious by complementation. Deletions (of nucleotides 417, 999, and 1017) mapped in the L proteinase and capsid protein-coding regions. Cell killing followed two-hit kinetics, defective genomes were encapsidated into separate viral particles, and individual viral plaques contained defective genomes with no detectable standard FMDV RNA. Infection in the absence of standard FMDV RNA was achieved by cotransfection of susceptible cells with transcripts produced in vitro from plasmids encoding the defective genomes. These results document the first step of an evolutionary transition toward genome segmentation of an unsegmented RNA virus and provide an experimental system to compare rates of RNA progeny production and resistance to enhanced mutagenesis of a segmented genome versus its unsegmented counterpart.
Figures
References
-
- Alberts, B., A. Johnson, J. Lewis, M. Raff, K. Roberts, and P. Walter. 2002. Molecular biology of the cell. Garland Science, New York, N.Y.
-
- Baranowski, E., C. M. Ruiz-Jarabo, and E. Domingo. 2001. Evolution of cell recognition by viruses. Science 292:1102-1105. - PubMed
-
- Chao, L. 1991. Levels of selection, evolution of sex in RNA viruses, and the origin of life. J. Theor. Biol. 153:229-246. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
