

Genetic steps of mammalian homologous repair with distinct mutagenic consequences
- PMID: 15485900
- PMCID: PMC522275
- DOI: 10.1128/MCB.24.21.9305-9316.2004
Genetic steps of mammalian homologous repair with distinct mutagenic consequences
Abstract
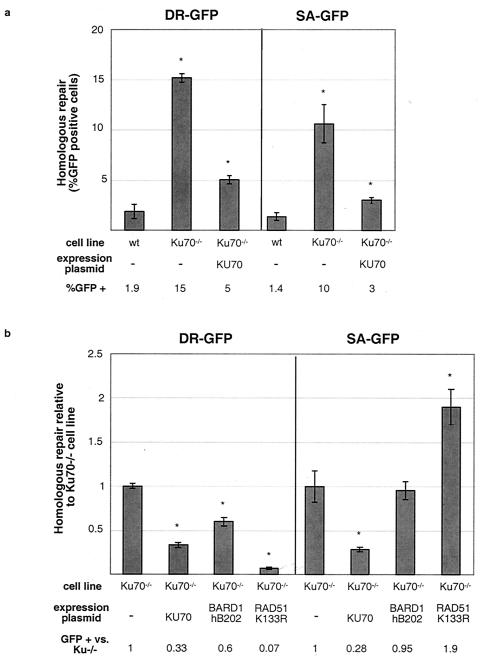
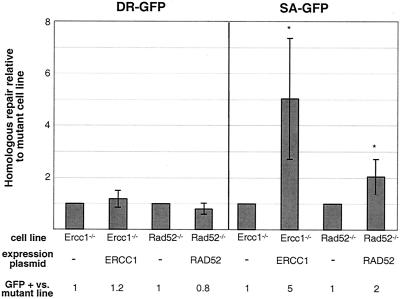
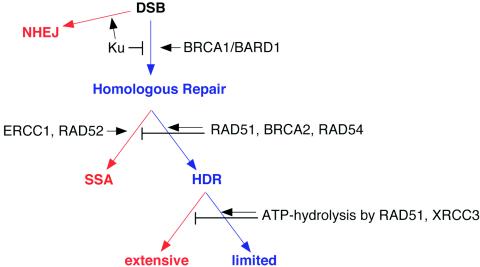
Repair of chromosomal breaks is essential for cellular viability, but misrepair generates mutations and gross chromosomal rearrangements. We investigated the interrelationship between two homologous-repair pathways, i.e., mutagenic single-strand annealing (SSA) and precise homology-directed repair (HDR). For this, we analyzed the efficiency of repair in mammalian cells in which double-strand break (DSB) repair components were disrupted. We observed an inverse relationship between HDR and SSA when RAD51 or BRCA2 was impaired, i.e., HDR was reduced but SSA was increased. In particular, expression of an ATP-binding mutant of RAD51 led to a >90-fold shift to mutagenic SSA repair. Additionally, we found that expression of an ATP hydrolysis mutant of RAD51 resulted in more extensive gene conversion, which increases genetic loss during HDR. Disruption of two other DSB repair components affected both SSA and HDR, but in opposite directions: SSA and HDR were reduced by mutation of Brca1, which, like Brca2, predisposes to breast cancer, whereas SSA and HDR were increased by Ku70 mutation, which affects nonhomologous end joining. Disruption of the BRCA1-associated protein BARD1 had effects similar to those of mutation of BRCA1. Thus, BRCA1/BARD1 has a role in homologous repair before the branch point of HDR and SSA. Interestingly, we found that Ku70 mutation partially suppresses the homologous-repair defects of BARD1 disruption. We also examined the role of RAD52 in homologous repair. In contrast to yeast, Rad52(-)(/)(-) mouse cells had no detectable HDR defect, although SSA was decreased. These results imply that the proper genetic interplay of repair factors is essential to limit the mutagenic potential of DSB repair.
Figures
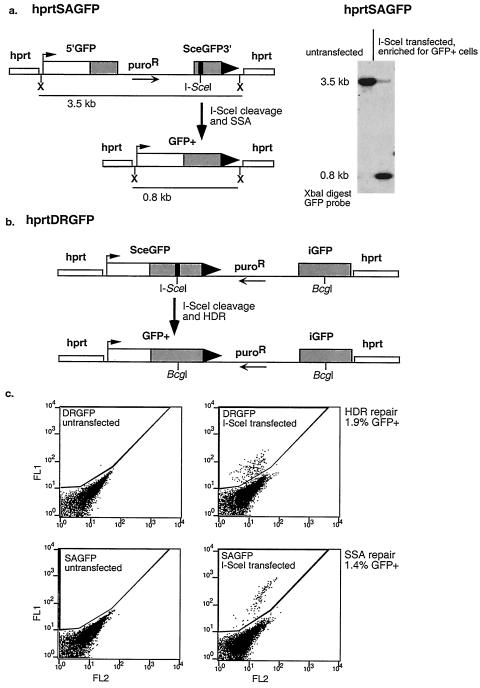
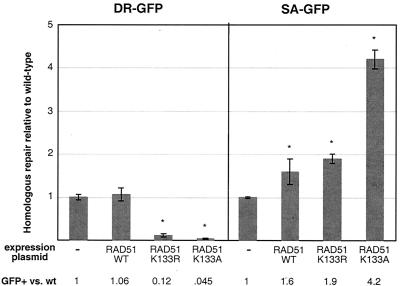
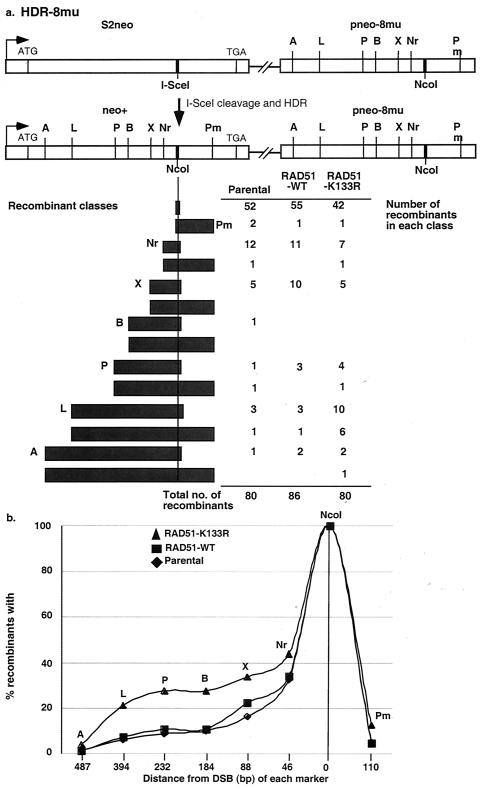
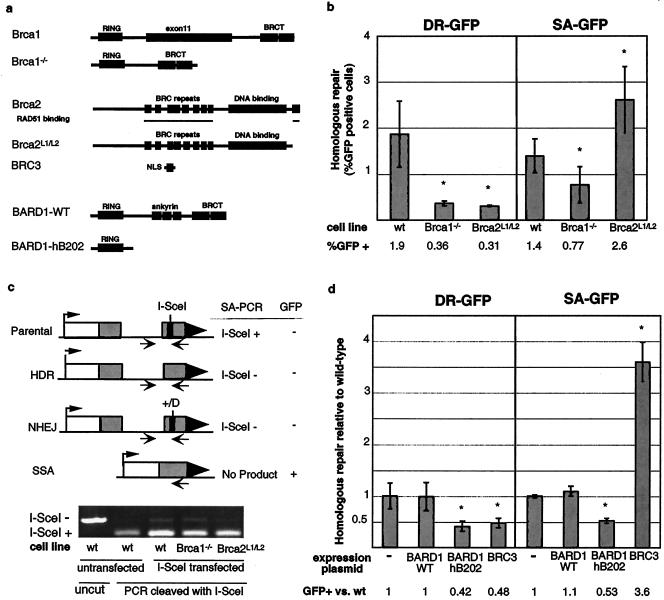
References
-
- Adams, M. D., M. McVey, and J. J. Sekelsky. 2003. Drosophila BLM in double-strand break repair by synthesis-dependent strand annealing. Science 299:265-267. - PubMed
-
- Albertson, D. G., C. Collins, F. McCormick, and J. W. Gray. 2003. Chromosome aberrations in solid tumors. Nat. Genet. 34:369-376. - PubMed
-
- Brenneman, M. A., B. M. Wagener, C. A. Miller, C. Allen, and J. A. Nickoloff. 2002. XRCC3 controls the fidelity of homologous recombination: roles for XRCC3 in late stages of recombination. Mol. Cell 10:387-395. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous