

Identification of the proteins required for biosynthesis of diphthamide, the target of bacterial ADP-ribosylating toxins on translation elongation factor 2
- PMID: 15485916
- PMCID: PMC522255
- DOI: 10.1128/MCB.24.21.9487-9497.2004
Identification of the proteins required for biosynthesis of diphthamide, the target of bacterial ADP-ribosylating toxins on translation elongation factor 2
Abstract
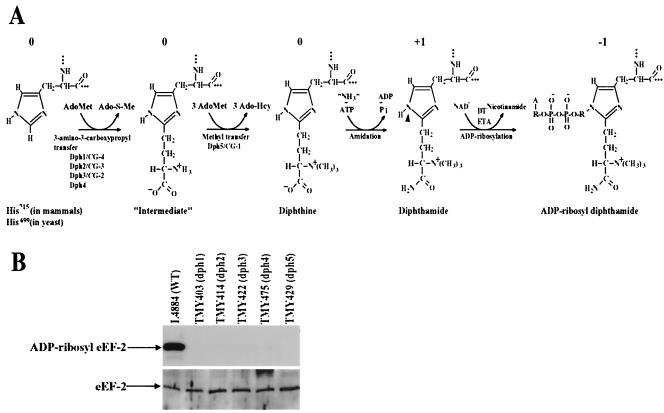
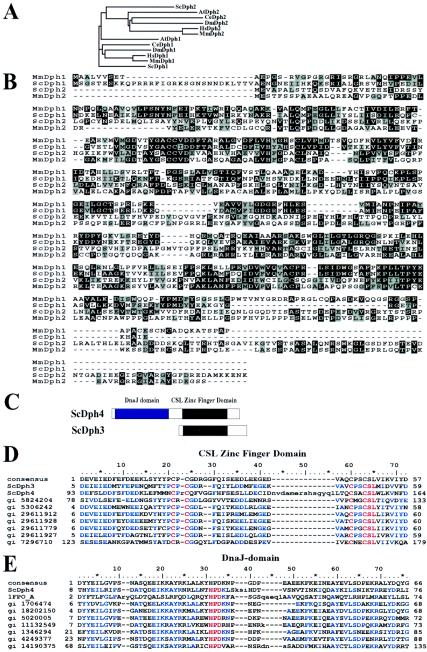
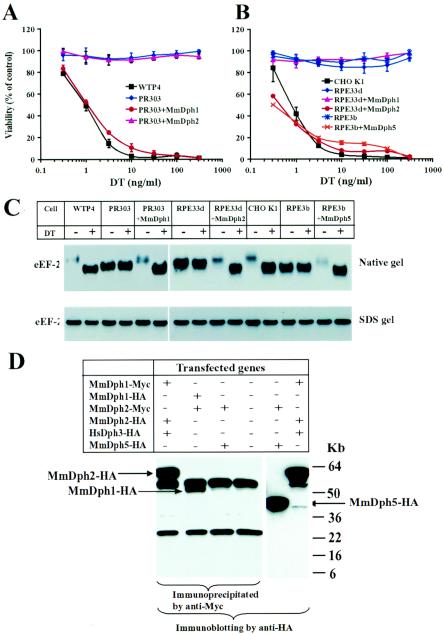
Diphthamide, a posttranslational modification of translation elongation factor 2 that is conserved in all eukaryotes and archaebacteria and is the target of diphtheria toxin, is formed in yeast by the actions of five proteins, Dph1 to -5, and a still unidentified amidating enzyme. Dph2 and Dph5 were previously identified. Here, we report the identification of the remaining three yeast proteins (Dph1, -3, and -4) and show that all five Dph proteins have either functional (Dph1, -2, -3, and -5) or sequence (Dph4) homologs in mammals. We propose a unified nomenclature for these proteins (e.g., HsDph1 to -5 for the human proteins) and their genes based on the yeast nomenclature. We show that Dph1 and Dph2 are homologous in sequence but functionally independent. The human tumor suppressor gene OVCA1, previously identified as homologous to yeast DPH2, is shown to actually be HsDPH1. We show that HsDPH3 is the previously described human diphtheria toxin and Pseudomonas exotoxin A sensitivity required gene 1 and that DPH4 encodes a CSL zinc finger-containing DnaJ-like protein. Other features of these genes are also discussed. The physiological function of diphthamide and the basis of its ubiquity remain a mystery, but evidence is presented that Dph1 to -3 function in vivo as a protein complex in multiple cellular processes.
Figures
References
-
- Bruening, W., A. H. Prowse, D. C. Schultz, M. Holgado-Madruga, A. Wong, and A. K. Godwin. 1999. Expression of OVCA1, a candidate tumor suppressor, is reduced in tumors and inhibits growth of ovarian cancer cells. Cancer Res. 59:4973-4983. - PubMed
-
- Burns, N., B. Grimwade, P. B. Ross-Macdonald, E. Y. Choi, K. Finberg, G. S. Roeder, and M. Snyder. 1994. Large-scale analysis of gene expression, protein localization, and gene disruption in Saccharomyces cerevisiae. Genes Dev. 8:1087-1105. - PubMed
-
- Carroll, S. F., J. T. Barbieri, and R. J. Collier. 1988. Diphtheria toxin: purification and properties. Methods Enzymol. 165:68-76. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases