

A novel molecular recognition motif necessary for targeting photoactivated phytochrome signaling to specific basic helix-loop-helix transcription factors
- PMID: 15486100
- PMCID: PMC527196
- DOI: 10.1105/tpc.104.025643
A novel molecular recognition motif necessary for targeting photoactivated phytochrome signaling to specific basic helix-loop-helix transcription factors
Abstract
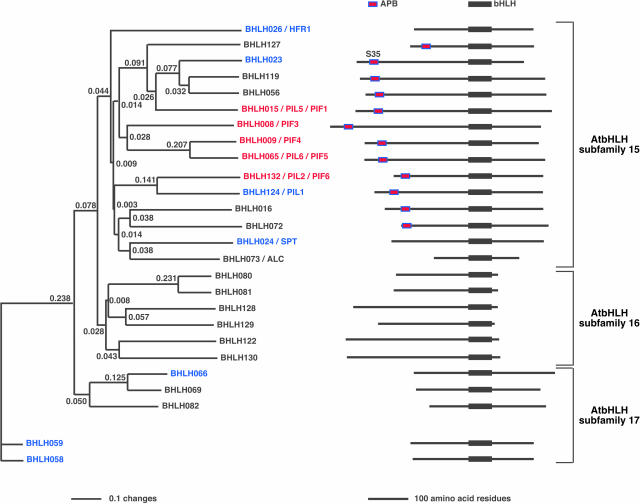
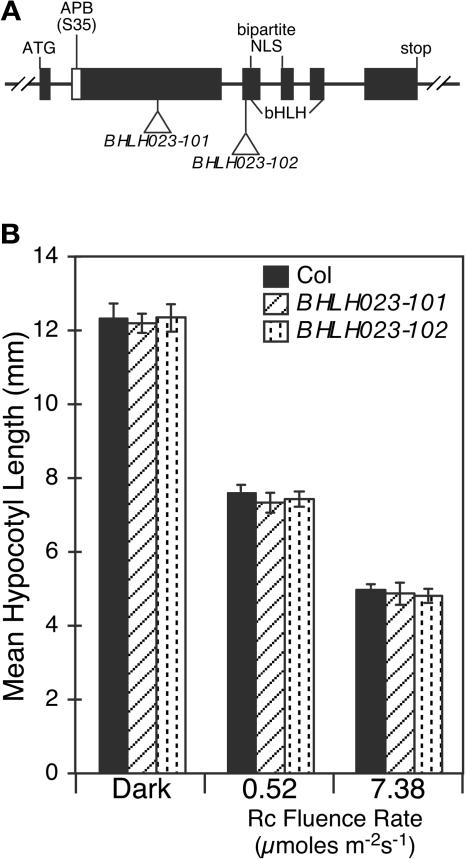
The phytochrome (phy) family of sensory photoreceptors (phyA to phyE) in Arabidopsis thaliana control plant developmental transitions in response to informational light signals throughout the life cycle. The photoactivated conformer of the photoreceptor Pfr has been shown to translocate into the nucleus where it induces changes in gene expression by an unknown mechanism. Here, we have identified two basic helix-loop-helix (bHLH) transcription factors, designated PHYTOCHROME-INTERACTING FACTOR5 (PIF5) and PIF6, which interact specifically with the Pfr form of phyB. These two factors cluster tightly with PIF3 and two other phy-interacting bHLH proteins in a phylogenetic subfamily within the large Arabidopsis bHLH (AtbHLH) family. We have identified a novel sequence motif (designated the active phytochrome binding [APB] motif) that is conserved in these phy-interacting AtbHLHs but not in other noninteractors. Using the isolated domain and site-directed mutagenesis, we have shown that this motif is both necessary and sufficient for binding to phyB. Transgenic expression of the native APB-containing AtbHLH protein, PIF4, in a pif4 null mutant, rescued the photoresponse defect in this mutant, whereas mutated PIF4 constructs with site-directed substitutions in conserved APB residues did not. These data indicate that the APB motif is necessary for PIF4 function in light-regulated seedling development and suggest that conformer-specific binding of phyB to PIF4 via the APB motif is necessary for this function in vivo. Binding assays with the isolated APB domain detected interaction with phyB, but none of the other four Arabidopsis phys. Collectively, the data suggest that the APB domain provides a phyB-specific recognition module within the AtbHLH family, thereby conferring photoreceptor target specificity on a subset of these transcription factors and, thus, the potential for selective signal channeling to segments of the transcriptional network.
Figures






Similar articles
-
PIF4, a phytochrome-interacting bHLH factor, functions as a negative regulator of phytochrome B signaling in Arabidopsis.EMBO J. 2002 May 15;21(10):2441-50. doi: 10.1093/emboj/21.10.2441. EMBO J. 2002. PMID: 12006496 Free PMC article.
-
Phytochrome B binds with greater apparent affinity than phytochrome A to the basic helix-loop-helix factor PIF3 in a reaction requiring the PAS domain of PIF3.Proc Natl Acad Sci U S A. 2000 Nov 21;97(24):13419-24. doi: 10.1073/pnas.230433797. Proc Natl Acad Sci U S A. 2000. PMID: 11069292 Free PMC article.
-
PIL5, a phytochrome-interacting basic helix-loop-helix protein, is a key negative regulator of seed germination in Arabidopsis thaliana.Plant Cell. 2004 Nov;16(11):3045-58. doi: 10.1105/tpc.104.025163. Epub 2004 Oct 14. Plant Cell. 2004. PMID: 15486102 Free PMC article.
-
bHLH class transcription factors take centre stage in phytochrome signalling.Trends Plant Sci. 2005 Feb;10(2):51-4. doi: 10.1016/j.tplants.2004.12.005. Trends Plant Sci. 2005. PMID: 15708340 Review.
-
Phytochrome Interacting Factors: central players in phytochrome-mediated light signaling networks.Trends Plant Sci. 2007 Nov;12(11):514-521. doi: 10.1016/j.tplants.2007.10.001. Epub 2007 Oct 22. Trends Plant Sci. 2007. PMID: 17933576 Review.
Cited by
-
The Transcriptional Regulator BBX19 Promotes Hypocotyl Growth by Facilitating COP1-Mediated EARLY FLOWERING3 Degradation in Arabidopsis.Plant Cell. 2015 Apr;27(4):1128-39. doi: 10.1105/tpc.15.00044. Epub 2015 Apr 3. Plant Cell. 2015. PMID: 25841036 Free PMC article.
-
The phytochrome-interacting vascular plant one-zinc finger1 and VOZ2 redundantly regulate flowering in Arabidopsis.Plant Cell. 2012 Aug;24(8):3248-63. doi: 10.1105/tpc.112.101915. Epub 2012 Aug 17. Plant Cell. 2012. PMID: 22904146 Free PMC article.
-
Phytochrome Interacting Factors (PIFs) in Solanum lycopersicum: Diversity, Evolutionary History and Expression Profiling during Different Developmental Processes.PLoS One. 2016 Nov 1;11(11):e0165929. doi: 10.1371/journal.pone.0165929. eCollection 2016. PLoS One. 2016. PMID: 27802334 Free PMC article.
-
KIDARI, encoding a non-DNA Binding bHLH protein, represses light signal transduction in Arabidopsis thaliana.Plant Mol Biol. 2006 May;61(1-2):283-96. doi: 10.1007/s11103-006-0010-2. Plant Mol Biol. 2006. PMID: 16786307
-
Photoperiod Control of Plant Growth: Flowering Time Genes Beyond Flowering.Front Plant Sci. 2022 Feb 9;12:805635. doi: 10.3389/fpls.2021.805635. eCollection 2021. Front Plant Sci. 2022. PMID: 35222453 Free PMC article. Review.
References
-
- Bauer, D., Viczián, A., Kircher, S., Nobis, T., Nitschke, R., Kunkel, T., Panigrahi, K., Ádám, E., Fejes, E., Schäfer, E., and Nagy, F. (2004). COP1 and multiple photoreceptors control degradation of PIF3, a transcription factor, required for light signalling in Arabidopsis. Plant Cell 16, 1433–1445. - PMC - PubMed
-
- Clough, S.J., and Bent, A.F. (1998). Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
