

Genome-wide mRNA surveillance is coupled to mRNA export
- PMID: 15489286
- PMCID: PMC525545
- DOI: 10.1101/gad.1241204
Genome-wide mRNA surveillance is coupled to mRNA export
Abstract
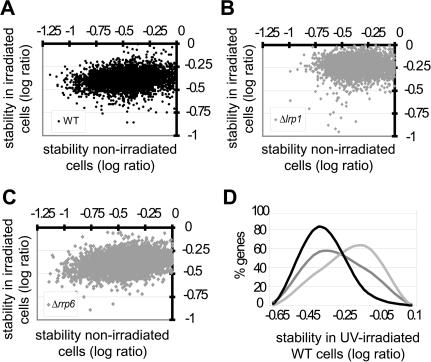
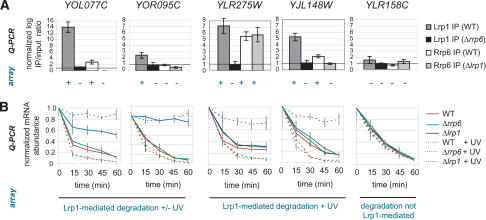
Nuclear export of mRNA is a central step in gene expression that shows extensive coupling to transcription and transcript processing. However, little is known about the fate of mRNA and its export under conditions that damage the DNA template and RNA itself. Here we report the discovery of four new factors required for mRNA export through a screen of all annotated nonessential Saccharomyces cerevisiae genes. Two of these factors, mRNA surveillance factor Rrp6 and DNA repair protein Lrp1, are nuclear exosome components that physically interact with one another. We find that Lrp1 mediates specific mRNA degradation upon DNA-damaging UV irradiation as well as general mRNA degradation. Lrp1 requires Rrp6 for genomic localization to genes encoding its mRNA targets, and Rrp6 genomic localization in turn correlates with transcription. Further, Rrp6 and Lrp1 are both required for repair of UV-induced DNA damage. These results demonstrate coupling of mRNA surveillance to mRNA export and suggest specificity of the RNA surveillance machinery for different transcript populations. Broadly, these findings link DNA and RNA surveillance to mRNA export.
Figures



References
-
- Amberg D.C., Goldstein, A.L., and Cole, C.N. 1992. Isolation and characterization of RAT1: An essential gene of Saccharomyces cerevisiae required for the efficient nucleocytoplasmic trafficking of mRNA. Genes & Dev. 6: 1173-1189. - PubMed
-
- Andrulis E.D., Werner, J., Nazarian, A., Erdjument-Bromage, H., Tempst, P., and Lis, J.T. 2002. The RNA processing exosome is linked to elongating RNA polymerase II in Drosophila. Nature 420: 837-841. - PubMed
-
- Aoki K., Suzuki, K., Sugano, T., Tasaka, T., Nakahara, K., Kuge, O., Omori, A., and Kasai, M. 1995. A novel gene, Translin, encodes a recombination hotspot binding protein associated with chromosomal translocations. Nat. Genet. 10: 167-174. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous