

Comparative Study
doi: 10.1101/gad.317004.
The active spatial organization of the beta-globin locus requires the transcription factor EKLF
Affiliations
- PMID: 15489291
- PMCID: PMC529536
- DOI: 10.1101/gad.317004
Item in Clipboard
Comparative Study
The active spatial organization of the beta-globin locus requires the transcription factor EKLF
Genes Dev.
.
Abstract
Three-dimensional organization of a gene locus is important for its regulation, as recently demonstrated for the beta-globin locus. When actively expressed, the cis-regulatory elements of the beta-globin locus are in proximity in the nuclear space, forming a compartment termed the Active Chromatin Hub (ACH). However, it is unknown which proteins are involved in ACH formation. Here, we show that EKLF, an erythroid transcription factor required for adult beta-globin gene transcription, is also required for ACH formation. We conclude that transcription factors can play an essential role in the three-dimensional organization of gene loci.
Figures
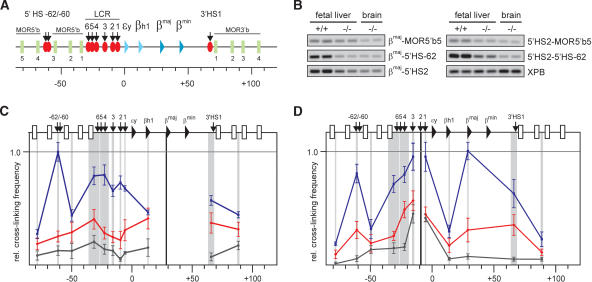
EKLF influences the spatial organization of the β-globin locus. (A) Schematic presentation of the mouse β-globin locus. Globin genes are indicated by triangles: light blue for embryonic and dark blue for the fetal/adult genes. Olfactory receptor genes (MOR5′b and MOR3′b) are indicated by green rectangles and are numbered. DNaseI HS are shown as red ovals with arrows. The scale is in kilobases. (B) Examples of PCR-amplified ligation products run on a 2% agarose gel. Primer combinations are shown on the right. XPB is used to standardize the amount of template (Palstra et al. 2003). (+/+) Wild type; (-/-) EKLF knockout. (C,D) Locus-wide relative cross-linking frequencies in E12.5 fetal livers. Results obtained with wild-type livers are shown in blue, EKLF-/- livers in red, nonexpressing brains in black. The X axis shows position in the locus. Gray shading indicates the positions and sizes of the HindIII fragments containing primers used in the PCR analysis. Black shading represents the position of the fragment containing the fixed primer in the HindIII fragment of the βmaj-gene (C) or 5′HS2 (D). Within each graph, the highest cross-linking frequency value is set to 1. Error bars indicate S.E.M.
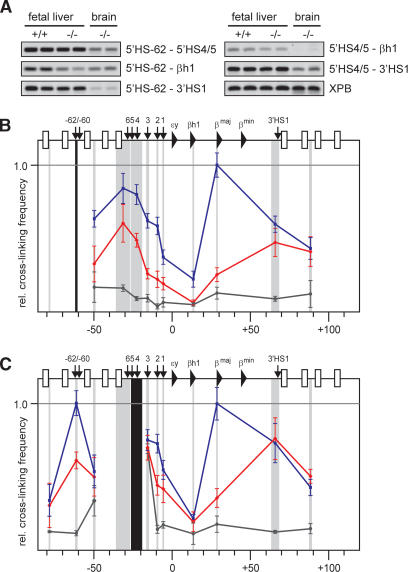
An ACH substructure is formed independent of EKLF. (A) Examples of PCR-amplified ligation products run on a 2% agarose gel. Primer combinations are shown on the right. (B,C) Locus-wide relative cross-linking frequencies of HindIII restriction fragments containing 5′HS-62 (b), and 5′HS4/5 (c). See the legend for Figure 1 for other details.
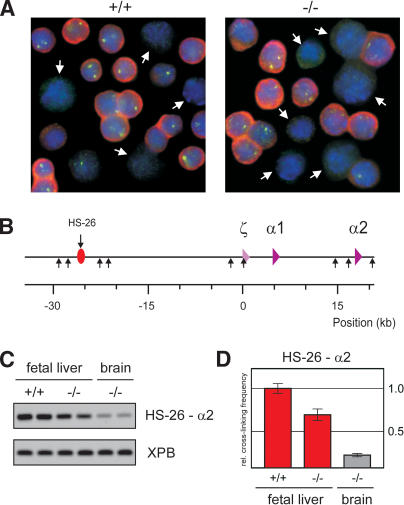
HS-26-promoter interactions in the α-globin locus are not affected by EKLF. (A) In situ hybridization of E12.5 fetal liver cells of wild-type and EKLF-/- fetuses, detecting α-globin mRNA (red) and primary transcripts (green). DAPI staining (blue) is used to show nuclear DNA. White arrows indicate cells that were scored negative for α-globin expression. (B) Schematic drawing of the mouse α-globin locus. The red oval with arrow depicts the position of the HS-26 distal regulatory element. The α-like globin genes are indicated by purple triangles. (Small arrows) HindIII restriction sites. (C) Example of PCR-amplified ligation products of HindIII restriction fragments containing HS-26 and α2 in E12.5 fetal liver and brain cells of wild-type and EKLF-/- fetuses. The XPB PCR product is used as template control. (D) Quantified data of PCR-amplified ligation products. (Red) Fetal liver; (gray) brain. Error bars indicate S.E.M. The cross-linking frequency in wild-type fetal liver cells is set to one.
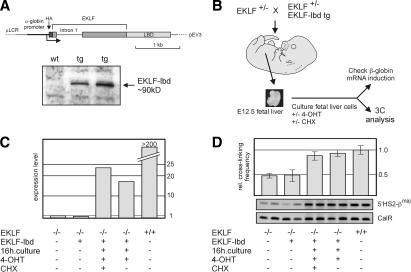
EKLF is directly involved in the spatial organization of the β-globin locus. (A) Schematic drawing of the EKLF–lbd expression construct used to generate transgenic mice. The Western blot shows expression of the EKLF–lbd fusion protein in the fetal livers of transgenic mice detected by an antibody recognizing the HA tag. (B) Flow chart of the experimental design. Fetal livers are isolated from E12.5 control and EKLF null::EKLF–lbd tg fetuses and disrupted, and the erythroid cells are cultured in the presence of 4-OHT with or without CHX for 16 h. Cells are then harvested, cross-linked with formaldehyde, and subjected to 3C analysis. From a portion of the cells, RNA is isolated to check β-globin gene expression. (C) Expression of β-globin analyzed by real-time RT–PCR. Expression of Hprt was used to standardize the β-globin expression levels. Representative experiment is shown. (D) 3C analysis of the interactions between 5′HS2 and the β-globin promoter. Representative examples of the PCR reactions are shown. Error bars indicate S.E.M. Calreticulin was used as template control.
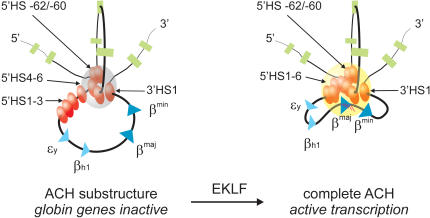
The formation of the complete ACH requires the presence of EKLF. A two-dimensional representation of the proposed three-dimensional structure of the ACH is shown. The ACH is a nuclear compartment dedicated to RNA polymerase II transcription, formed by cis-regulatory elements of the β-globin locus (Palstra et al. 2003). In erythroid cells, a substructure of the ACH, consisting of 5′HS-62/-60, 3′HS1 and HS at the 5′ side of the LCR, is formed independently of EKLF. Progression of this substructure to a fully functional ACH, including the HS at the 3′ side of the LCR and the active β-globin gene, is dependent on the presence of EKLF. (Gray sphere) ACH substructure; (yellow sphere) ACH. RNA transcripts are indicated as red lines. See the legend for Figure 1A for other details.
References
-
- Armstrong J.A., Bieker, J.J., and Emerson, B.M. 1998. A SWI/SNF-related chromatin remodeling complex, E-RC1, is required for tissue-specific transcriptional regulation by EKLF in vitro. Cell 95: 93-104. - PubMed
-
- Bender M.A., Bulger, M., Close, J., and Groudine, M. 2000. β-globin gene switching and DNase I sensitivity of the endogenous β-globin locus in mice do not require the locus control region. Mol. Cell 5: 387-393. - PubMed
-
- Bieker J.J. 2001. Kruppel-like factors: Three fingers in many pies. J. Biol. Chem. 276: 34355-34358. - PubMed
-
- Carter D., Chakalova, L., Osborne, C.S., Dai, Y.F., and Fraser, P. 2002. Long-range chromatin regulatory interactions in vivo. Nat. Genet. 32: 623-626. - PubMed
-
- De Laat W. and Grosveld, F. 2003. Spatial organization of gene expression: The Active Chromatin Hub. Chromosome Res. 5: 447-459. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases