

The C-terminal alphaO helix of human Ogg1 is essential for 8-oxoguanine DNA glycosylase activity: the mitochondrial beta-Ogg1 lacks this domain and does not have glycosylase activity
- PMID: 15494448
- PMCID: PMC524278
- DOI: 10.1093/nar/gkh863
The C-terminal alphaO helix of human Ogg1 is essential for 8-oxoguanine DNA glycosylase activity: the mitochondrial beta-Ogg1 lacks this domain and does not have glycosylase activity
Abstract
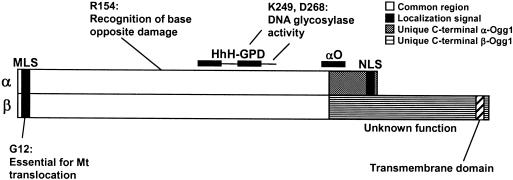
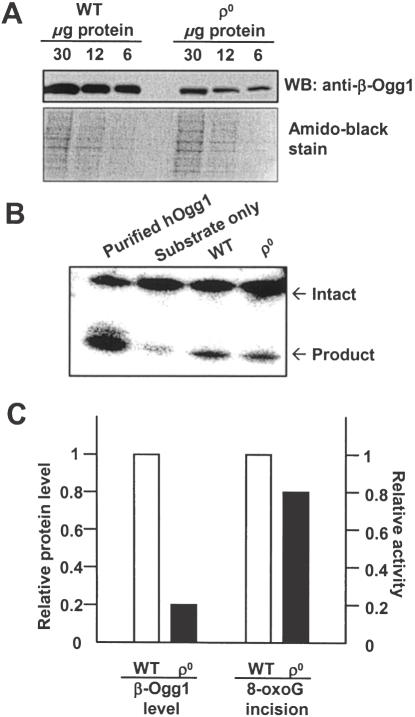
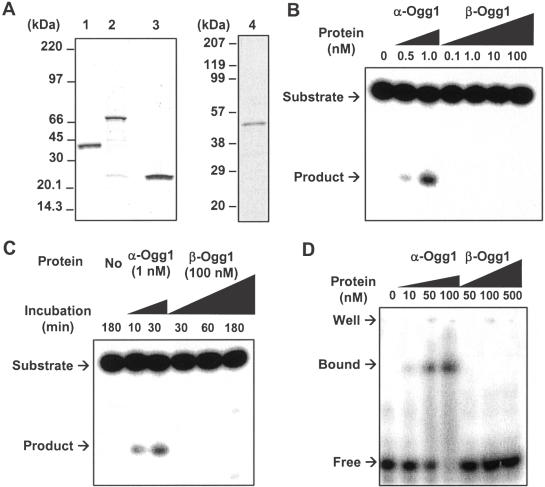
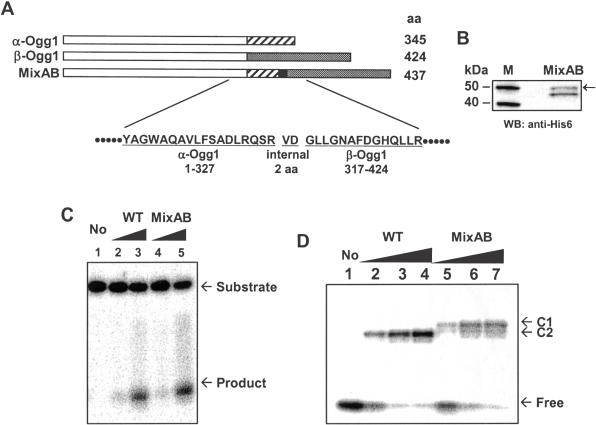
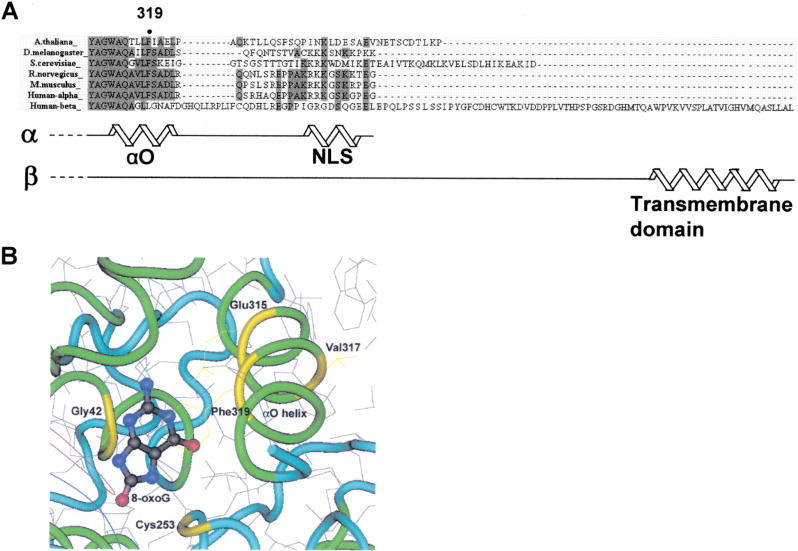
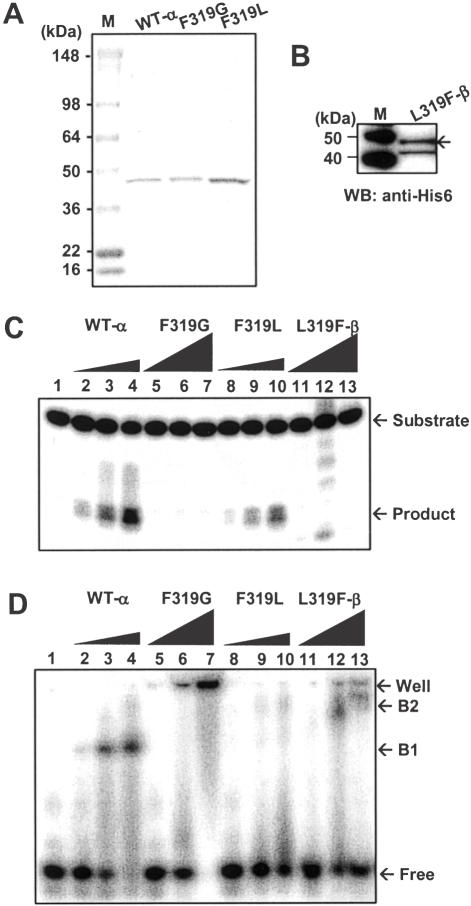
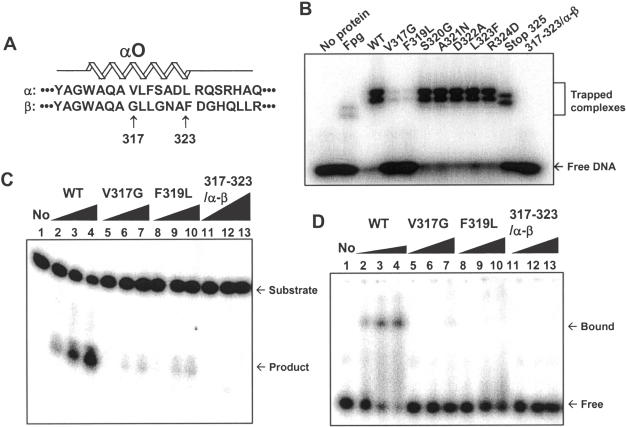
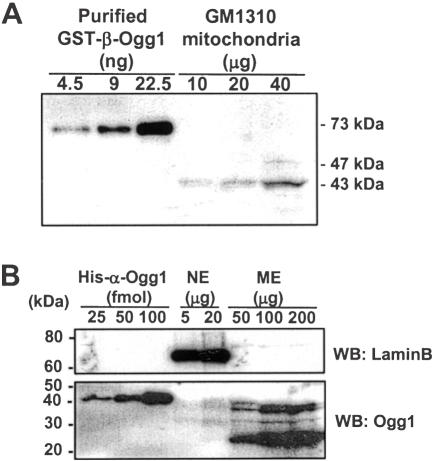
The human Ogg1 glycosylase is responsible for repairing 8-oxo-7,8-dihydroguanine (8-oxoG) in both nuclear and mitochondrial DNA. Two distinct Ogg1 isoforms are present; alpha-Ogg1, which mainly localizes to the nucleus and beta-Ogg1, which localizes only to mitochondria. We recently showed that mitochondria from rho(0) cells, which lack mitochondrial DNA, have similar 8-oxoG DNA glycosylase activity to that of wild-type cells. Here, we show that beta-Ogg1 protein levels are approximately 80% reduced in rho(0) cells, suggesting beta-Ogg1 is not responsible for 8-oxoG incision in mitochondria. Thus, we characterized the biochemical properties of recombinant beta-Ogg1. Surprisingly, recombinant beta-Ogg1 did not show any significant 8-oxoG DNA glycosylase activity in vitro. Since beta-Ogg1 lacks the C-terminal alphaO helix present in alpha-Ogg1, we generated mutant proteins with various amino acid substitutions in this domain. Of the seven amino acid positions substituted (317-323), we identified Val-317 as a novel critical residue for 8-oxoG binding and incision. Our results suggest that the alphaO helix is absolutely necessary for 8-oxoG DNA glycosylase activity, and thus its absence may explain why beta-Ogg1 does not catalyze 8-oxoG incision in vitro. Western blot analysis revealed the presence of significant amounts of alpha-Ogg1 in human mitochondria. Together with previous localization studies in vivo, this suggests that alpha-Ogg1 protein may provide the 8-oxoG DNA glycosylase activity for the repair of these lesions in human mitochondrial DNA. beta-Ogg1 may play a novel role in human mitochondria.
Figures
Similar articles
-
Cloning and characterization of an ascidian homolog of the human 8-oxoguanine DNA glycosylase (Ogg1) that is involved in the repair of 8-oxo-7,8-dihydroguanine in DNA in Ciona intestinalis.Int J Radiat Biol. 2006 Apr;82(4):241-50. doi: 10.1080/09553000600649216. Int J Radiat Biol. 2006. PMID: 16690592
-
Repair of 8-oxodeoxyguanosine lesions in mitochondrial dna depends on the oxoguanine dna glycosylase (OGG1) gene and 8-oxoguanine accumulates in the mitochondrial dna of OGG1-defective mice.Cancer Res. 2001 Jul 15;61(14):5378-81. Cancer Res. 2001. PMID: 11454679
-
Enzyme kinetics of an alternative splicing isoform of mitochondrial 8-oxoguanine DNA glycosylase, ogg1-1b, and compared with the nuclear ogg1-1a.J Biochem Mol Toxicol. 2015 Feb;29(2):49-56. doi: 10.1002/jbt.21605. Epub 2014 Sep 4. J Biochem Mol Toxicol. 2015. PMID: 25196052
-
Repair of 8-oxo-7,8-dihydroguanine in prokaryotic and eukaryotic cells: Properties and biological roles of the Fpg and OGG1 DNA N-glycosylases.Free Radic Biol Med. 2017 Jun;107:179-201. doi: 10.1016/j.freeradbiomed.2016.11.042. Epub 2016 Nov 27. Free Radic Biol Med. 2017. PMID: 27903453 Review.
-
An active alternative splicing isoform of human mitochondrial 8-oxoguanine DNA glycosylase (OGG1).Genes Environ. 2015 Oct 1;37:21. doi: 10.1186/s41021-015-0021-9. eCollection 2015. Genes Environ. 2015. PMID: 27350816 Free PMC article. Review.
Cited by
-
From cell senescence to age-related diseases: differential mechanisms of action of senescence-associated secretory phenotypes.BMB Rep. 2015 Oct;48(10):549-58. doi: 10.5483/bmbrep.2015.48.10.122. BMB Rep. 2015. PMID: 26129674 Free PMC article. Review.
-
Human 8-oxoguanine glycosylase OGG1 binds nucleosome at the dsDNA ends and the super-helical locations.Commun Biol. 2024 Sep 28;7(1):1202. doi: 10.1038/s42003-024-06919-7. Commun Biol. 2024. PMID: 39341999 Free PMC article.
-
DNA damage and repair: relevance to mechanisms of neurodegeneration.J Neuropathol Exp Neurol. 2008 May;67(5):377-87. doi: 10.1097/NEN.0b013e31816ff780. J Neuropathol Exp Neurol. 2008. PMID: 18431258 Free PMC article. Review.
-
Mitochondrial DNA, base excision repair and neurodegeneration.DNA Repair (Amst). 2008 Jul 1;7(7):1098-109. doi: 10.1016/j.dnarep.2008.03.011. Epub 2008 May 16. DNA Repair (Amst). 2008. PMID: 18485834 Free PMC article. Review.
-
Mitochondrial base excision repair assays.Methods. 2010 Aug;51(4):416-25. doi: 10.1016/j.ymeth.2010.02.020. Epub 2010 Feb 25. Methods. 2010. PMID: 20188838 Free PMC article. Review.
References
-
- Sies H. (1997) Oxidative stress: oxidants and antioxidants. Exp. Physiol., 82, 291–295. - PubMed
-
- Dizdaroglu M., Jaruga,P., Birincioglu,M. and Rodriguez,H. (2002) Free radical-induced damage to DNA: mechanisms and measurement. Free Radic. Biol. Med., 32, 1102–1115. - PubMed
-
- Souza-Pinto N.C. and Bohr,V.A. (2002) The mitochondrial theory of aging: involvement of mitochondrial DNA damage and repair. Int. Rev. Neurobiol., 53, 519–534. - PubMed
-
- Migliore L. and Coppede,F. (2002) Genetic and environmental factors in cancer and neurodegenerative diseases. Mutat. Res., 512, 135–153. - PubMed
-
- Shibutani S., Takeshita,M. and Grollman,A.P. (1991) Insertion of specific bases during DNA synthesis past the oxidation-damaged base 8-oxodG. Nature, 349, 431–434. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
