

The 'window' T-type calcium current in brain dynamics of different behavioural states
- PMID: 15498803
- PMCID: PMC1665496
- DOI: 10.1113/jphysiol.2004.076273
The 'window' T-type calcium current in brain dynamics of different behavioural states
Abstract
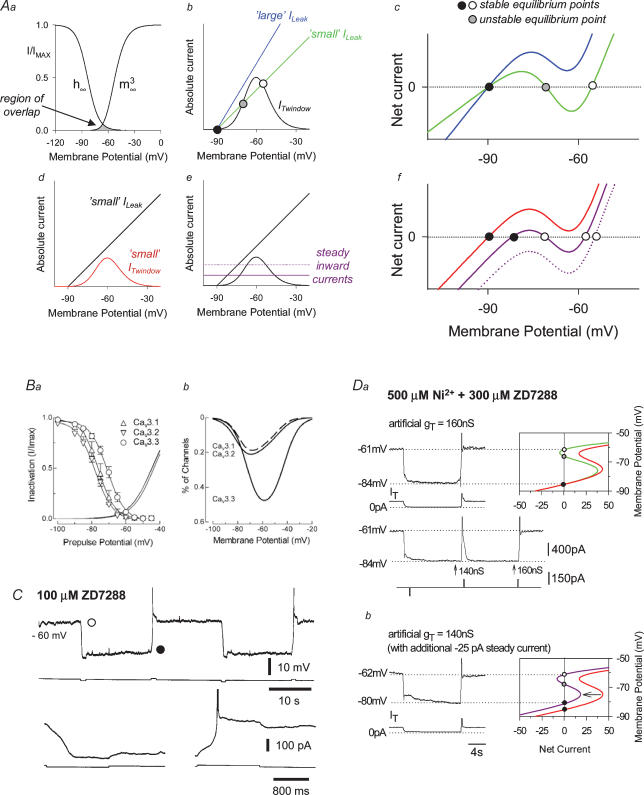
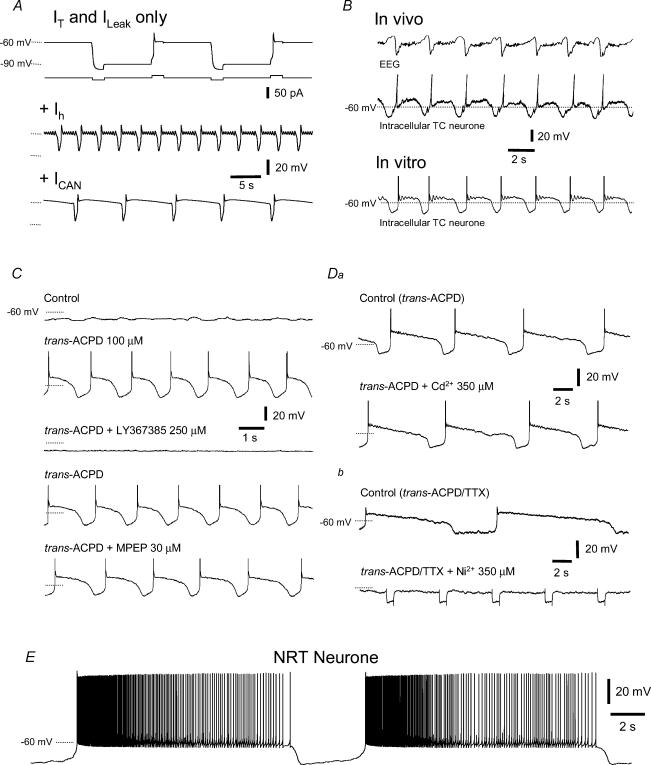
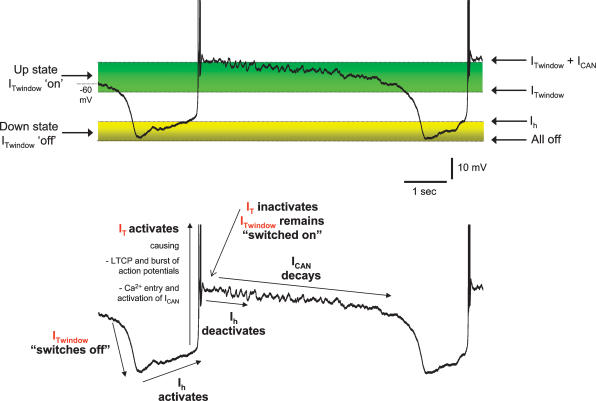
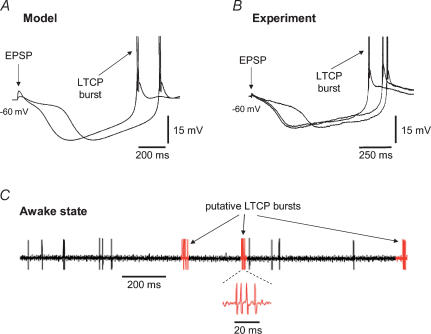
All three forms of recombinant low voltage-activated T-type Ca(2)(+) channels (Ca(v)3.1, Ca(v)3.2 and Ca(v)3.3) exhibit a small, though clearly evident, window T-type Ca(2)(+) current (I(Twindow)) which is also present in native channels from different neuronal types. In thalamocortical (TC) and nucleus reticularis thalami (NRT) neurones, and possibly in neocortical cells, an I(Twindow)-mediated bistability is the key cellular mechanism underlying the expression of the slow (< 1 Hz) sleep oscillation, one of the fundamental EEG rhythms of non-REM sleep. As the I(Twindow)-mediated bistability may also represent one of the cellular mechanisms underlying the expression of high frequency burst firing in awake conditions, I(Twindow) is of critical importance in neuronal population dynamics associated with different behavioural states.
Figures
References
-
- Achermann P, Borbely AA. Low-frequency (< 1 Hz) oscillations in the human sleep electroencephalogram. Neuroscience. 1997;81:213–222. - PubMed
-
- Amzica F, Steriade M. The K-complex: its slow (<1-Hz) rhythmicity and relation to delta waves. Neurology. 1997;49:952–959. - PubMed
-
- Blethyn KL, Hughes SW, Cope DW, Crunelli V. Nucleus-specific properties of the slow (<1 hz) oscillation in thalamic neurones in vitro. Soc Neurosci Abstr. 2002;28(352.4)
-
- Blethyn KL, Hughes SW, Cope DW, Crunelli V. The role of ionic conductances underlying a slow (<1 hz) oscillation in neurones of the thalamic reticular nucleus in vitro. Soc Neurosci Abstr. 2003;29(699.3)
-
- Carbone E, Lux HD. A low voltage-activated, fully inactivating Ca channel in vertebrate sensory neurones. Nature. 1984;310:501–502. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous