

Uterine spiral artery remodeling involves endothelial apoptosis induced by extravillous trophoblasts through Fas/FasL interactions
- PMID: 15499040
- PMCID: PMC4228192
- DOI: 10.1161/01.ATV.0000148547.70187.89
Uterine spiral artery remodeling involves endothelial apoptosis induced by extravillous trophoblasts through Fas/FasL interactions
Abstract
Objective: Invasion of uterine spiral arteries by extravillous trophoblasts in the first trimester of pregnancy results in loss of endothelial and musculoelastic layers. This remodeling is crucial for an adequate blood supply to the fetus with a failure to remodel implicated in the etiology of the hypertensive disorder preeclampsia. The mechanism by which trophoblasts induce this key process is unknown. This study gives the first insights into the potential mechanisms involved.
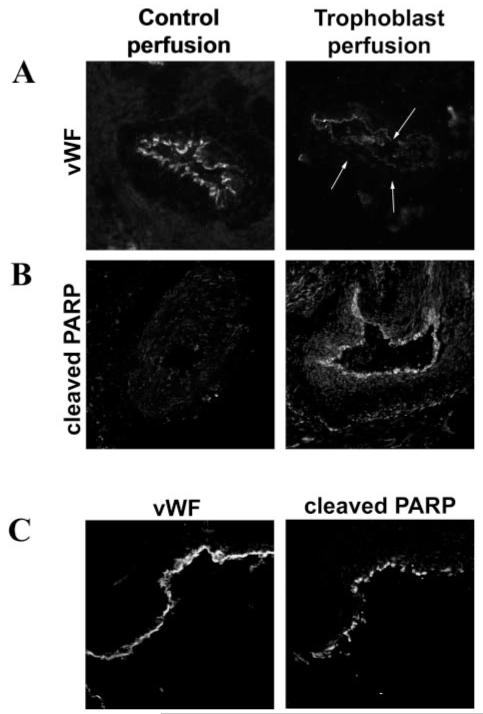
Methods and results: Spiral arteries were dissected from nonplacental bed biopsies obtained at Caesarean section, and a novel model was used to mimic in vivo events. Arteries were cultured with trophoblasts in the lumen, and apoptotic changes in the endothelial layer were detected after 20 hours, leading to loss of endothelium by 96 hours. In vitro, coculture experiments showed that trophoblasts stimulated apoptosis of primary decidual endothelial cells and an endothelial cell line. This was blocked by caspase inhibition and NOK2, a FasL blocking antibody. NOK2 also abrogated trophoblast-induced endothelial apoptosis in the vessel model.
Conclusions: Extravillous trophoblast induction of endothelial apoptosis is a possible mechanism by which the endothelium is removed, and vascular remodeling may occur in uterine spiral arteries. Fas/FasL interactions have an important role in trophoblast-induced endothelial apoptosis.
Figures





References
-
- Pijnenborg R, Dixon G, Robertson WB, Brosens I. Trophoblastic invasion of human decidua from 8 to 18 weeks of pregnancy. Placenta. 1980;1:3–19. - PubMed
-
- Enders AC, Blankenship TN. Modification of endometrial arteries during invasion by cytotrophoblast cells in the pregnant macaque. Acta Anat. 1997;159:169–193. - PubMed
-
- Brosens JJ, Pijnenborg R, Brosens IA. The myometrial junctional zone spiral arteries in normal and abnormal pregnancies: a review of the literature. Am J Obstet Gynecol. 2002;187:1416–1423. - PubMed
-
- Hung TH, Skepper JN, Charnock-Jones DS, Burton GJ. Hypoxia-reoxygenation: a potent inducer of apoptotic changes in the human placenta and possible etiological factor in preeclampsia. Circ Res. 2002;90:1274–1281. - PubMed
-
- Barker DJ. The long-term outcome of retarded fetal growth. Clin Obstet Gynecol. 1997;40:853–863. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
