

Glycosylation of the major polar tube protein of Encephalitozoon hellem, a microsporidian parasite that infects humans
- PMID: 15501763
- PMCID: PMC523040
- DOI: 10.1128/IAI.72.11.6341-6350.2004
Glycosylation of the major polar tube protein of Encephalitozoon hellem, a microsporidian parasite that infects humans
Abstract
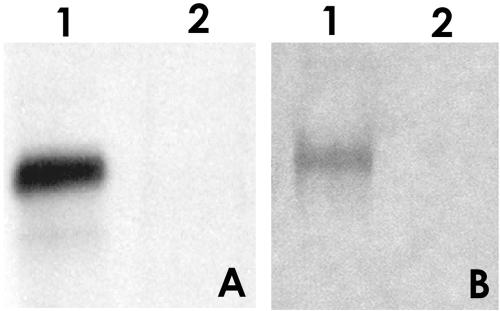
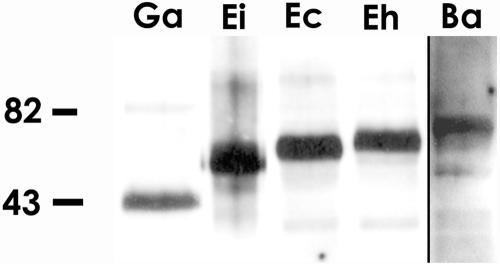
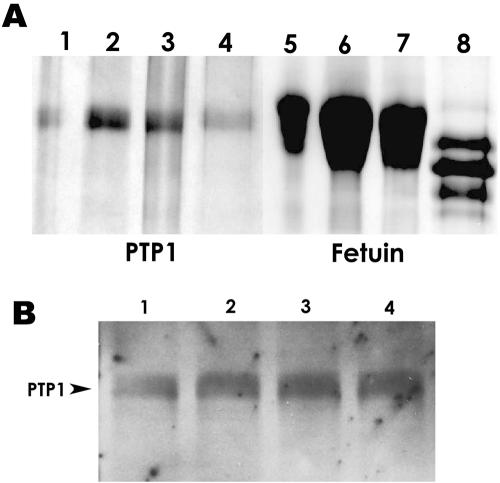
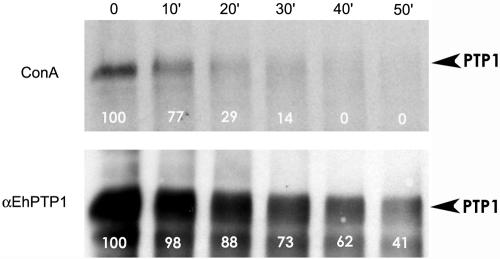
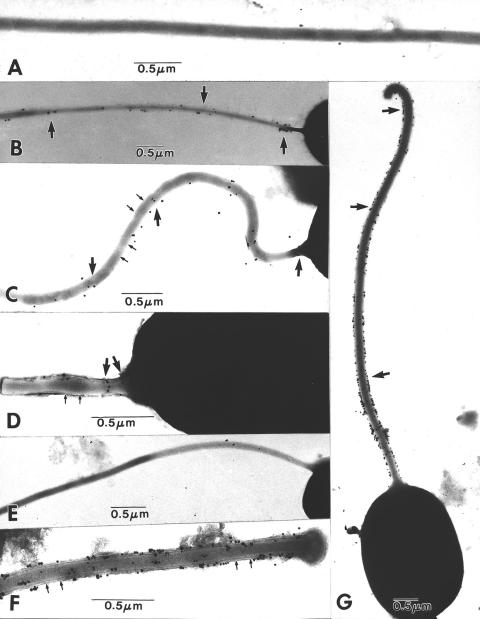
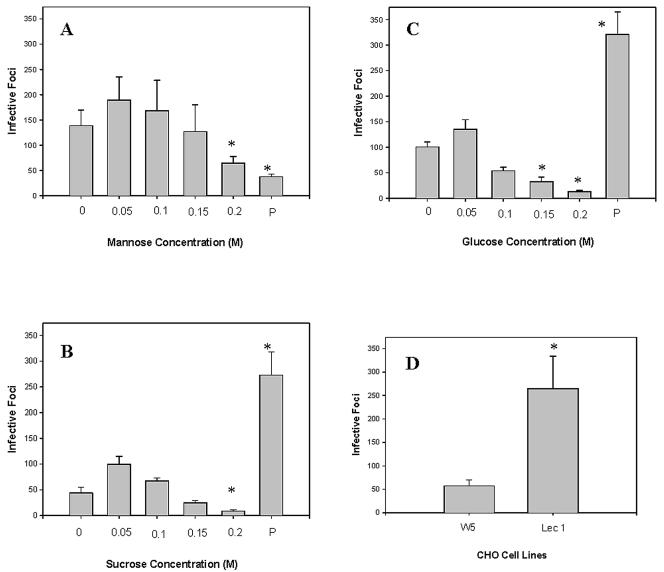
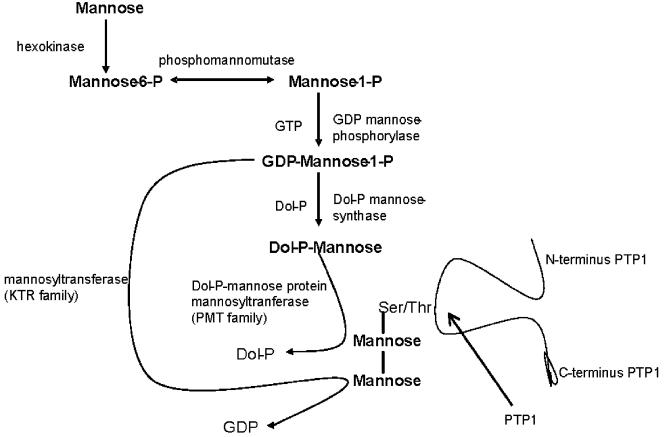
The microsporidia are ubiquitous, obligate intracellular eukaryotic spore-forming parasites infecting a wide range of invertebrates and vertebrates, including humans. The defining structure of microsporidia is the polar tube, which forms a hollow tube through which the sporoplasm is transferred to the host cell. Research on the molecular and cellular biology of the polar tube has resulted in the identification of three polar tube proteins: PTP1, PTP2, and PTP3. The major polar tube protein, PTP1, accounts for at least 70% of the mass of the polar tube. In the present study, PTP1 was found to be posttranslationally modified. Concanavalin A (ConA) bound to PTP1 and to the polar tube of several different microsporidia species. Analysis of the glycosylation of Encephalitozoon hellem PTP1 suggested that it is modified by O-linked mannosylation, and ConA binds to these O-linked mannose residues. Mannose pretreatment of RK13 host cells decreased their infection by E. hellem, consistent with an interaction between the mannosylation of PTP1 and some unknown host cell mannose-binding molecule. A CHO cell line (Lec1) that is unable to synthesize complex-type N-linked oligosaccharides had an increased susceptibility to E. hellem infection compared to wild-type CHO cells. These data suggest that the O-mannosylation of PTP1 may have functional significance for the ability of microsporidia to invade their host cells.
Figures

References
-
- Aronson, M., O. Medalia, L. Schori, D. Mirelman, N. Sharon, and I. Ofek. 1979. Prevention of colonization of the urinary tract of mice with Escherichia coli by blocking of bacterial adherence with methyl alpha-D-mannopyranoside. J. Infect. Dis. 139:329-332. - PubMed
-
- Cali, A., L. M. Weiss, and P. M. Takvorian. 2002. Brachiola algerae spore membrane systems, their activity during extrusion, and a new structural entity, the multilayered interlaced network, associated with the polar tube and the sporoplasm. J. Eukaryot. Microbiol. 49:164-174. - PubMed
-
- Chen, X. M., and N. F. LaRusso. 2000. Mechanisms of attachment and internalization of Cryptosporidium parvum to biliary and intestinal epithelial cells. Gastroenterology 118:368-379. - PubMed
-
- Delbac, F., P. Peyret, G. Metenier, D. David, A. Danchin, and C. P. Vivares. 1998. On proteins of the microsporidian invasive apparatus: complete sequence of a polar tube protein of Encephalitozoon cuniculi. Mol. Microbiol. 29:825-834. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
