

The Type II heat-labile enterotoxins LT-IIa and LT-IIb and their respective B pentamers differentially induce and regulate cytokine production in human monocytic cells
- PMID: 15501764
- PMCID: PMC523043
- DOI: 10.1128/IAI.72.11.6351-6358.2004
The Type II heat-labile enterotoxins LT-IIa and LT-IIb and their respective B pentamers differentially induce and regulate cytokine production in human monocytic cells
Abstract
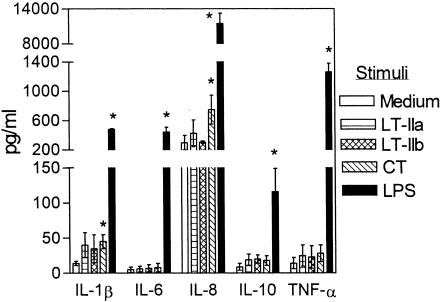
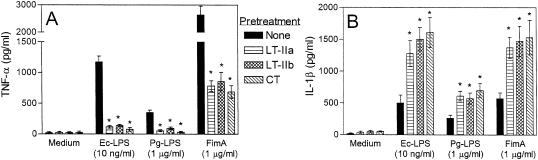
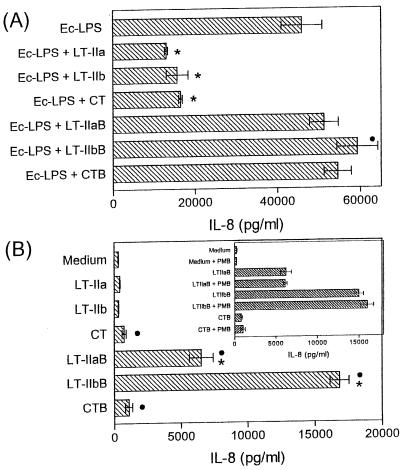
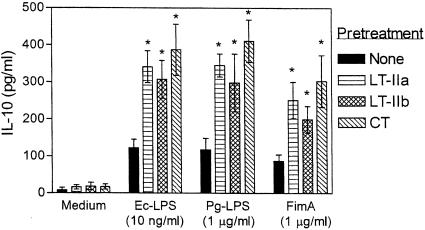
The type II heat-labile enterotoxins, LT-IIa and LT-IIb, exhibit potent adjuvant properties. However, little is known about their immunomodulatory activities upon interaction with innate immune cells, unlike the widely studied type I enterotoxins that include cholera toxin (CT). We therefore investigated interactions of LT-IIa and LT-IIb with human monocytic THP-1 cells. We found that LT-II enterotoxins were inactive in stimulating cytokine release, whereas CT induced low levels of interleukin-1beta (IL-1beta) and IL-8. However, all three enterotoxins potently regulated cytokine induction in cells activated by bacterial lipopolysaccharide or fimbriae. Induction of proinflammatory (tumor necrosis factor alpha [TNF-alpha]) or chemotactic (IL-8) cytokines was downregulated, whereas induction of cytokines with anti-inflammatory (IL-10) or mucosal adjuvant properties (IL-1beta) was upregulated by the enterotoxins. These effects appeared to depend on their A subunits, because isolated B-pentameric subunits lacked regulatory activity. Enterotoxin-mediated inhibition of proinflammatory cytokine induction in activated cells was partially attributable to synergism for endogenous production of IL-10 and to an IL-10-independent inhibition of nuclear factor kappaB (NF-kappaB) activation. In sharp contrast to the holotoxins, the B pentamers (LT-IIaB and, to a greater extent, LT-IIbB) stimulated cytokine production, suggesting a link between the absence of the A subunit and increased proinflammatory properties. In this regard, the ability of LT-IIbB to activate NF-kappaB and induce TNF-alpha and IL-8 was antagonized by the LT-IIb holotoxin. These findings support distinct immunomodulatory roles for the LT-II holotoxins and their respective B pentamers. Moreover, the anti-inflammatory properties of the holotoxins may serve to suppress innate immunity and promote the survival of the pathogen.
Figures


References
-
- Akira, S. 2001. Toll-like receptors and innate immunity. Adv. Immunol. 78:1-56. - PubMed
-
- Auwerx, J. 1991. The human leukemia cell line, THP-1: a multifaceted model for the study of monocyte-macrophage differentiation. Experientia 47:22-31. - PubMed
-
- Brenner, S., S. Prösch, K. Schenke-Layland, U. Riese, U. Gausmann, and C. Platzer. 2003. cAMP-induced interleukin-10 promoter activation depends on CCAAT/enhancer-binding protein expression and monocytic differentiation. J. Biol. Chem. 278:5597-5604. - PubMed
-
- Bromander, A., J. Holmgren, and N. Lycke. 1991. Cholera toxin stimulates IL-1 production and enhances antigen presentation by macrophages in vitro. J. Immunol. 146:2908-2914. - PubMed
-
- Chandra, G., J. Cogswell, L. Miller, M. Godlevski, S. Stinnett, S. Noel, S. Kadwell, T. Kost, and J. Gray. 1995. Cyclic AMP signaling pathways are important in IL-1 beta transcriptional regulation. J. Immunol. 155:4535-4543. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
