

Protein thiol modifications visualized in vivo
- PMID: 15502869
- PMCID: PMC521172
- DOI: 10.1371/journal.pbio.0020333
Protein thiol modifications visualized in vivo
Abstract
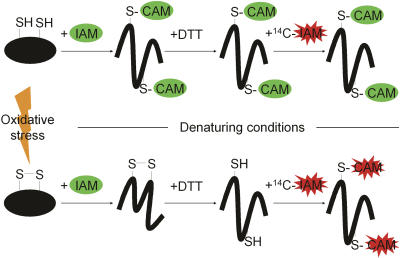
Thiol-disulfide interconversions play a crucial role in the chemistry of biological systems. They participate in the major systems that control the cellular redox potential and prevent oxidative damage. In addition, thiol-disulfide exchange reactions serve as molecular switches in a growing number of redox-regulated proteins. We developed a differential thiol-trapping technique combined with two-dimensional gel analysis, which in combination with genetic studies, allowed us to obtain a snapshot of the in vivo thiol status of cellular proteins. We determined the redox potential of protein thiols in vivo, identified and dissected the in vivo substrate proteins of the major cellular thiol-disulfide oxidoreductases, and discovered proteins that undergo thiol modifications during oxidative stress. Under normal growth conditions most cytosolic proteins had reduced cysteines, confirming existing dogmas. Among the few partly oxidized cytosolic proteins that we detected were proteins that are known to form disulfide bond intermediates transiently during their catalytic cycle (e.g., dihydrolipoyl transacetylase and lipoamide dehydrogenase). Most proteins with highly oxidized thiols were periplasmic proteins and were found to be in vivo substrates of the disulfide-bond-forming protein DsbA. We discovered a substantial number of redox-sensitive cytoplasmic proteins, whose thiol groups were significantly oxidized in strains lacking thioredoxin A. These included detoxifying enzymes as well as many metabolic enzymes with active-site cysteines that were not known to be substrates for thioredoxin. H(2)O(2)-induced oxidative stress resulted in the specific oxidation of thiols of proteins involved in detoxification of H(2)O(2) and of enzymes of cofactor and amino acid biosynthesis pathways such as thiolperoxidase, GTP-cyclohydrolase I, and the cobalamin-independent methionine synthase MetE. Remarkably, a number of these proteins were previously or are now shown to be redox regulated.
Conflict of interest statement
The authors have declared that no conflicts of interest exist.
Figures




References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
