

doi: 10.1073/pnas.0406844101.
Epub 2004 Oct 25.
Integration of reproductive meristem fates by a SEPALLATA-like MADS-box gene
Affiliations
- PMID: 15505223
- PMCID: PMC524820
- DOI: 10.1073/pnas.0406844101
Item in Clipboard
Integration of reproductive meristem fates by a SEPALLATA-like MADS-box gene
Proc Natl Acad Sci U S A.
.
Abstract
Reproductive transition, inflorescence architecture, meristem patterning, and floral organ identity have been studied as distinct research areas in plant science. By using the ornamental plant Gerbera, we demonstrate that all of these keystone aspects of reproductive meristematic fate are integrated genetically by a single SEPALLATA-like MADS-box gene from a functional class designated previously as "floral homeotic" or "organ identity." This extended regulatory network has not been elaborated in the model plant systems, which have a floral design and inflorescence-determinacy state that obscures these relationships.
Figures
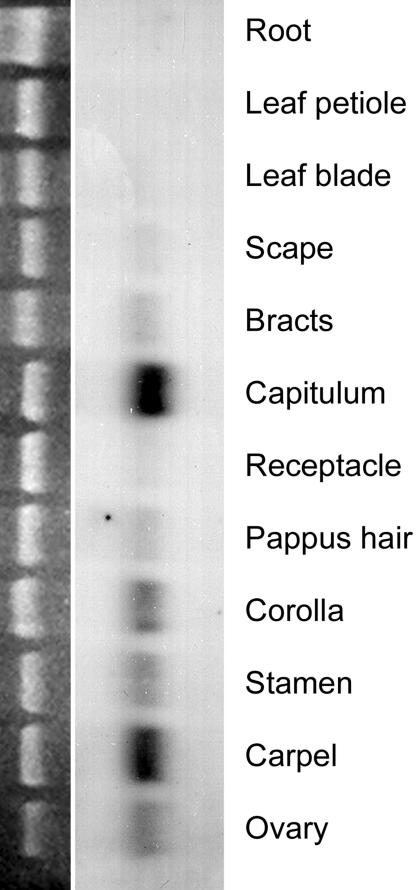
Expression of GRCD2 in various Gerbera tissues. (Right) Total RNA from different Gerbera organs, pooled from several developmental stages, was hybridized with GRCD2 full-length probe. The GRCD2 gene is expressed in all floral organs {pappus [i.e., sepals (14)], petals, stamens, and carpels} and other floral tissues, including ovary, bracts, scape, and receptacle. Expression was also detected in young capitula, suggesting that the GRCD2 gene is also active during early stages of inflorescence and flower development. Very weak expression was detected in the leaf blade, but there was no detectable expression in leaf petioles or roots. (Left) Equal loading of RNA was controlled with ethidium bromide staining (18S rRNA).
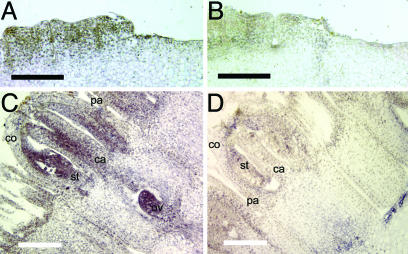
In situ hybridization analysis showing expression of GRCD2 in young Gerbera capitula. Longitudinal sections of wild-type capitulum [diameter, 8 mm (A and B) and 14 mm (C and D)] hybridized with antisense (A and C) and sense (B and D) GRCD2 probes. The initial expression of GRCD2 appears to be equal in all cells of young flower primordia from which flower organs have not yet emerged (A). When flowers reach the ring primordium stage (14), GRCD2 expression concentrates to the central part of the primordium, from which stamens and carpels will later emerge. In more mature flower primordia (C), in which the identities of floral organs have been determined, GRCD2 expression is strongest in stamens and carpels as well as in the developing ovule. In A and B, two flower primordia are visible. In C and D, a single flower primordium is shown, with pappus bristles (pa), corolla (co), stamens (st), carpels (ca), and the ovule (ov) indicated. [Scale bars indicate 200 μm (A and B) and 1 mm (C and D).]

Homeotic changes in transgenic Gerbera. (A) Disk flowers of non-transgenic Gerbera show pappus hairs in whorl 1 (w1), fused petals (corolla) in whorl 2 (w2), and fused anthers in whorl 3 (w3). The disk flower whorl 4 (w4) organs, carpels, are covered by the fused anthers in nontransgenic flowers. (B) In transgenic Gerbera in which GRCD2 is partially down regulated, green leaf-like organs replace carpels in w4. (C) In transgenic Gerbera in which GAGA1 is partially down regulated (14), w4 organs are similarly replaced by green structures, and concomitantly, w3 organs by petal-like structures. (D) Transgenic Gerbera with strongly reduced GRCD2 expression shows petal-like organs in w4. (E) Nontransgenic ray flower showing normal fertile carpels in w4. (Scale bars indicate 1 mm.)
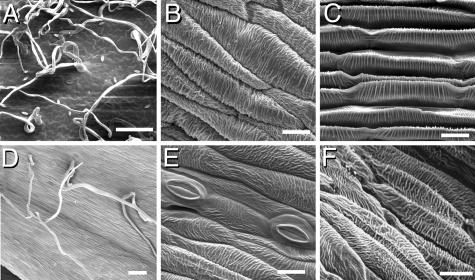
Epidermal characterization of the whorl 4 organs. SEM of the surface of a wild-type leaf (A), carpel (B), and petal (C). In GRCD2 down-regulated plants the whorl 4 organs show presence of multicellular hair-like structures (D) and stomata (E). Neither stomata nor hairs are present on normal carpels, but stomata are found on the abaxial surface of petals and, like hairs, on the adaxial and abaxial sides of leaves and bracts, as well as on the surfaces of scapes. In stronger phenotypes, the anti-GRCD2 whorl 4 surface (F) resembles the ridges typical to wild-type petals (C). The carpels of the transgenic plants were not functional, as confirmed by crossing tests with nontransgenic plants. [Scale bars indicate 100 μm (A and D) and 20 μm (B, C, E, and F).]
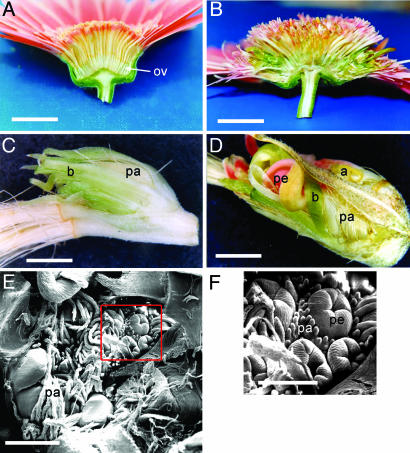
The ovaries in anti-GRCD2 Gerbera revert to inflorescences. (A) Wild-type inflorescence of Gerbera, which were cut in half to show inferior ovaries (ov) below the whorls of floral organs. (B) Transgenic Gerbera with reduced GRCD2 expression shows disrupted ovaries in which bracts (b), pappus hairs (pa), petals (pe), and anthers (a) develop instead of the ovule (C and D). (E and F) SEM micrograph of the inner meristematic regions replacing the ovary in anti-GRCD2 Gerbera shows several emerging flower primordia, each surrounded by its own pappus bristles (whorl 1) (F). [Scale bars indicate 2 cm (A and B), 2 mm (C and D), 500 μm (E), and 250 μm (F).]

Indeterminate growth of the inflorescence meristem in transgenic Gerbera. Development of wild-type (A, C, and E) and antisense GRCD2 (B, D, and F) inflorescence over time. The number of flower primordia is fixed at an early stage of development in wild-type Gerbera (A), whereas in the transgenic plants, the inflorescence meristem continues to proliferate (B). C and D show inflorescences at full flowering. At 35 days later, the wild-type inflorescence has matured and shed its seeds (E), whereas the transgenic inflorescences continue to produce flowers (F). The wild-type capitulum produces >600 florets (593 ± 56, n = 13), but the strong antisense GRCD2 lines produce up to 1,200 flowers (976 ± 98, n = 13). Transgenic plants in which GAGA2 was down-regulated (14) and that showed indeterminate flower meristems produced 594 ± 76 flowers (n = 6). The indeterminate growth mode of the anti-GRCD2 inflorescences was reflected both by continuously increasing production of disk flowers and by a concomitant enlargement of the receptacle. Although superficially unorganized, the spiral phyllotaxis of the transgenic inflorescences of the transgenic lines was not disturbed in terms of numbers of parastichies. [Scale bars indicate 1 mm (A and B) and 2 cm (C–F).]
References
-
- Battey, N. H. & Lyndon, R. F. (1990) Bot. Rev. 56, 162-189.
-
- Leyser, O. & Day, S. (2003) Mechanisms in Plant Development (Blackwell, Oxford).
-
- Pelaz, S., Ditta, G. S., Baumann, E., Wisman, E. & Yanofsky, M. F. (2000) Nature 405, 200-203. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
