

Guanosine tetra- and pentaphosphate synthase activity in chloroplasts of a higher plant: association with 70S ribosomes and inhibition by tetracycline
- PMID: 15507686
- PMCID: PMC528801
- DOI: 10.1093/nar/gkh916
Guanosine tetra- and pentaphosphate synthase activity in chloroplasts of a higher plant: association with 70S ribosomes and inhibition by tetracycline
Abstract
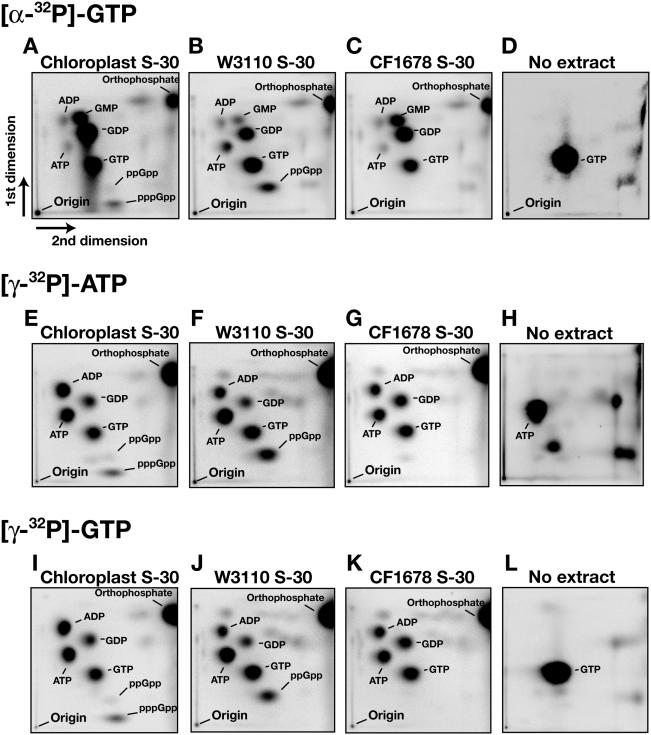
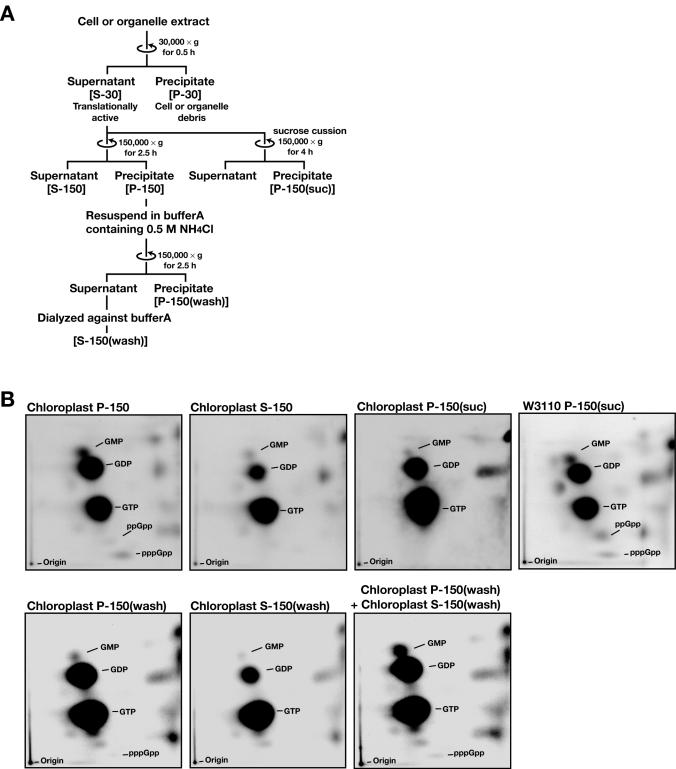
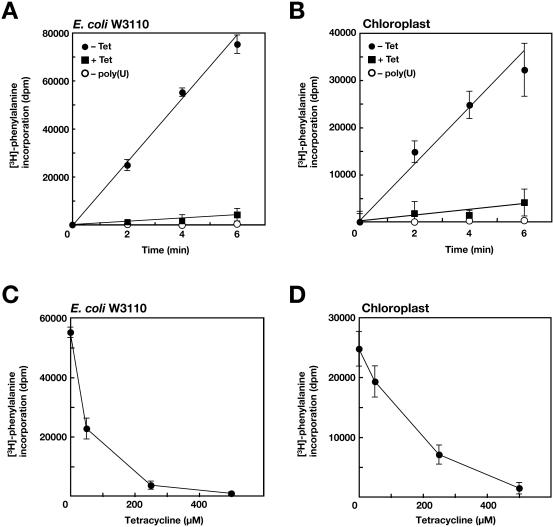
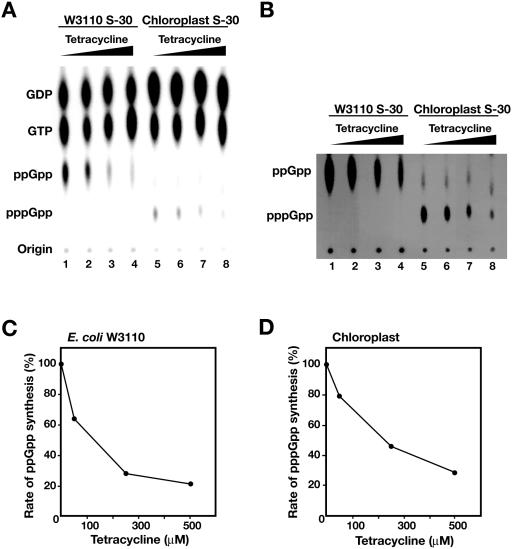
Chloroplasts possess bacterial-type systems for transcription and translation. On the basis of the identification of a Chlamydomonas reinhardtii gene encoding a RelA-SpoT homolog (RSH) that catalyzes the synthesis of guanosine tetra- or pentaphosphate [(p)ppGpp], we have previously suggested the operation of stringent control in the chloroplast genetic system. Although RSH genes have also been identified in several higher plants, the activities of the encoded enzymes and their mode of action in chloroplasts have remained uncharacterized. We have now characterized the intrinsic (p)ppGpp synthase activity of chloroplast extracts prepared from pea (Pisum sativum). Fractionation by ultracentrifugation suggested that the (p)ppGpp synthase activity of a translationally active chloroplast stromal extract was associated with 70S ribosomes. Furthermore, this enzymatic activity was inhibited by tetracycline, as was the peptide elongation activity of the extract. Structural comparisons between rRNA molecules of Escherichia coli and pea chloroplasts revealed the conservation of putative tetracycline-binding sites. These observations demonstrate the presence of a ribosome-associated (p)ppGpp synthase activity in the chloroplasts of a higher plant, further implicating (p)ppGpp in a genetic system of chloroplasts similar to that operative in bacteria.
Figures

Similar articles
-
A RelA-SpoT homolog (Cr-RSH) identified in Chlamydomonas reinhardtii generates stringent factor in vivo and localizes to chloroplasts in vitro.Nucleic Acids Res. 2002 Nov 15;30(22):4985-92. doi: 10.1093/nar/gkf628. Nucleic Acids Res. 2002. PMID: 12434003 Free PMC article.
-
Calcium-activated (p)ppGpp synthetase in chloroplasts of land plants.J Biol Chem. 2007 Dec 7;282(49):35536-45. doi: 10.1074/jbc.M703820200. Epub 2007 Oct 15. J Biol Chem. 2007. PMID: 17938177
-
Diversity in guanosine 3',5'-bisdiphosphate (ppGpp) sensitivity among guanylate kinases of bacteria and plants.J Biol Chem. 2014 May 30;289(22):15631-41. doi: 10.1074/jbc.M113.534768. Epub 2014 Apr 10. J Biol Chem. 2014. PMID: 24722991 Free PMC article.
-
Within and beyond the stringent response-RSH and (p)ppGpp in plants.Planta. 2017 Nov;246(5):817-842. doi: 10.1007/s00425-017-2780-y. Epub 2017 Sep 25. Planta. 2017. PMID: 28948393 Free PMC article. Review.
-
Signalling by the global regulatory molecule ppGpp in bacteria and chloroplasts of land plants.Plant Biol (Stuttg). 2011 Sep;13(5):699-709. doi: 10.1111/j.1438-8677.2011.00484.x. Epub 2011 May 31. Plant Biol (Stuttg). 2011. PMID: 21815973 Review.
Cited by
-
ppGpp inhibits peptide elongation cycle of chloroplast translation system in vitro.Plant Mol Biol. 2012 Jan;78(1-2):185-96. doi: 10.1007/s11103-011-9858-x. Epub 2011 Nov 23. Plant Mol Biol. 2012. PMID: 22108865
-
The Importance of Microorganisms for Sustainable Agriculture-A Review.Metabolites. 2022 Nov 11;12(11):1100. doi: 10.3390/metabo12111100. Metabolites. 2022. PMID: 36422239 Free PMC article. Review.
-
Multiple antibiotic resistance in Arabidopsis is conferred by mutations in a chloroplast-localized transport protein.Plant Physiol. 2009 Oct;151(2):559-73. doi: 10.1104/pp.109.143487. Epub 2009 Aug 12. Plant Physiol. 2009. PMID: 19675150 Free PMC article.
-
Antibiotics impact plant traits, even at small concentrations.AoB Plants. 2017 Mar 13;9(2):plx010. doi: 10.1093/aobpla/plx010. eCollection 2017 Mar. AoB Plants. 2017. PMID: 28439396 Free PMC article.
-
Antibiotic use and abuse: a threat to mitochondria and chloroplasts with impact on research, health, and environment.Bioessays. 2015 Oct;37(10):1045-53. doi: 10.1002/bies.201500071. Epub 2015 Sep 8. Bioessays. 2015. PMID: 26347282 Free PMC article.
References
-
- Cashel M., Gentry,D.R., Hernandez,V.J. and Vinella,D. (1996) Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd edn. ASM Press, Washington, DC, pp. 1458–1496.
-
- Chatterji D., Fujita,N. and Ishihama,A. (1998) The mediator for stringent control, ppGpp, binds to the β-subunit of Escherichia coli RNA polymerase. Genes Cells, 3, 279–287. - PubMed
-
- Toulokhonov I.I., Shulgina,I. and Hernandez,V.J. (2001) Binding of the transcription effector ppGpp to Escherichia coli RNA polymerase is allosteric, modular, and occurs near the N terminus of the β′-subunit. J. Biol. Chem., 276, 1220–1225. - PubMed
-
- Artsimovitch I., Patlan,V., Sekine,S., Vassylyeva,M.N., Hosaka,T., Ochi,K., Yokoyama,S. and Vassylyev,D. (2004) Structural basis for transcription regulation by alarmone ppGpp. Cell, 117, 299–310. - PubMed
-
- Sugiura M. (1992) The chloroplast genome. Plant Mol. Biol., 19, 149–168. - PubMed
