

Genetic and environmental factors influencing glutathione homeostasis in Saccharomyces cerevisiae
- PMID: 15509654
- PMCID: PMC539166
- DOI: 10.1091/mbc.e04-07-0560
Genetic and environmental factors influencing glutathione homeostasis in Saccharomyces cerevisiae
Abstract
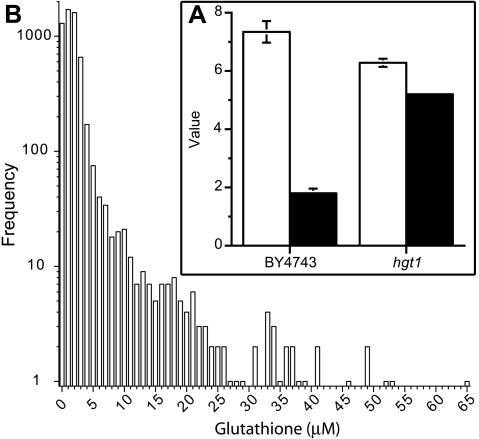
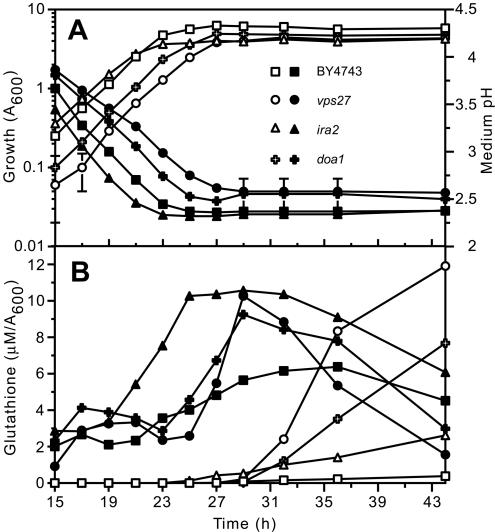
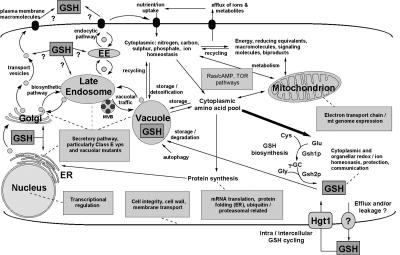
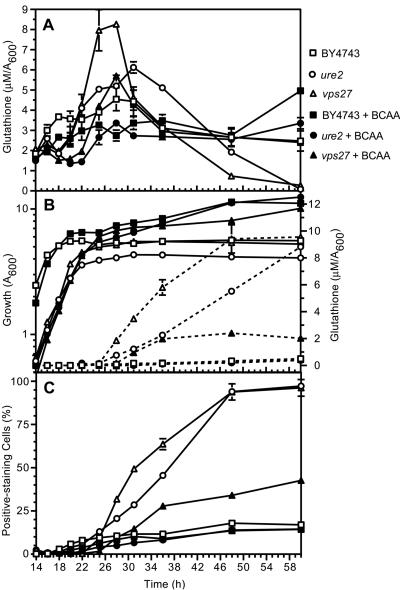
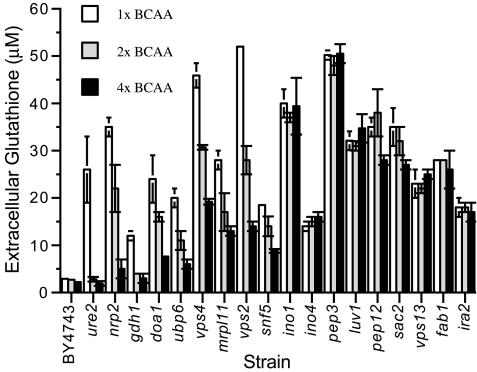
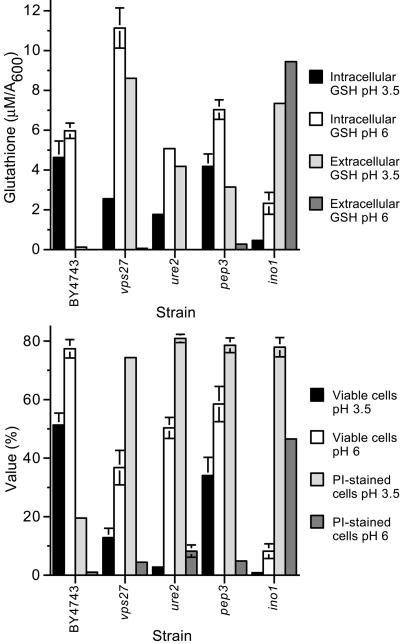
Glutathione is an essential metabolite protecting cells against oxidative stress and aging. Here, we show that endogenously synthesized glutathione undergoes intercellular cycling during growth to stationary phase. Genome-wide screening identified approximately 270 yeast deletion mutants that overexcrete glutathione, predominantly in the reduced form, and identified a surprising set of functions important for glutathione homeostasis. The highest excretors were affected in late endosome/vacuolar functions. Other functions identified included nitrogen/carbon source signaling, mitochondrial electron transport, ubiquitin/proteasomal processes, transcriptional regulation, ion transport and the cellular integrity pathway. For many mutants the availability of branched chain amino acids and extracellular pH influenced both glutathione homeostasis and cell viability. For all mutants tested, the onset of glutathione excretion occurred when intracellular concentration exceeded the maximal level found in the parental strain; however, in some mutants prolonged excretion led to substantial depletion of intracellular glutathione. These results significantly contribute to understanding mechanisms affecting glutathione homeostasis in eukaryotes and may provide insight into the underlying cause of glutathione depletion in degenerative processes such as Parkinson's disease. The important implications of these data for use of the yeast deletion collection for the study of other phenomena also are discussed.
Figures
Similar articles
-
Lactic-acid stress causes vacuolar fragmentation and impairs intracellular amino-acid homeostasis in Saccharomyces cerevisiae.J Biosci Bioeng. 2012 Apr;113(4):421-30. doi: 10.1016/j.jbiosc.2011.11.010. Epub 2011 Dec 15. J Biosci Bioeng. 2012. PMID: 22177309
-
Possible roles of vacuolar H+-ATPase and mitochondrial function in tolerance to air-drying stress revealed by genome-wide screening of Saccharomyces cerevisiae deletion strains.Yeast. 2008 Mar;25(3):179-90. doi: 10.1002/yea.1577. Yeast. 2008. PMID: 18224659
-
Vesicular Trafficking Systems Impact TORC1-Controlled Transcriptional Programs in Saccharomyces cerevisiae.G3 (Bethesda). 2016 Jan 6;6(3):641-52. doi: 10.1534/g3.115.023911. G3 (Bethesda). 2016. PMID: 26739646 Free PMC article.
-
Protein transport from the late Golgi to the vacuole in the yeast Saccharomyces cerevisiae.Biochim Biophys Acta. 2005 Jul 10;1744(3):438-54. doi: 10.1016/j.bbamcr.2005.04.004. Biochim Biophys Acta. 2005. PMID: 15913810 Review.
-
Regulation of Amino Acid Transport in Saccharomyces cerevisiae.Microbiol Mol Biol Rev. 2019 Oct 16;83(4):e00024-19. doi: 10.1128/MMBR.00024-19. Print 2019 Nov 20. Microbiol Mol Biol Rev. 2019. PMID: 31619504 Free PMC article. Review.
Cited by
-
Zap1 regulates zinc homeostasis and modulates virulence in Cryptococcus gattii.PLoS One. 2012;7(8):e43773. doi: 10.1371/journal.pone.0043773. Epub 2012 Aug 20. PLoS One. 2012. PMID: 22916306 Free PMC article.
-
Glutathione-Mediated Conjugation of Anticancer Drugs: An Overview of Reaction Mechanisms and Biological Significance for Drug Detoxification and Bioactivation.Molecules. 2022 Aug 17;27(16):5252. doi: 10.3390/molecules27165252. Molecules. 2022. PMID: 36014491 Free PMC article. Review.
-
An Optimized Competitive-Aging Method Reveals Gene-Drug Interactions Underlying the Chronological Lifespan of Saccharomyces cerevisiae.Front Genet. 2020 May 14;11:468. doi: 10.3389/fgene.2020.00468. eCollection 2020. Front Genet. 2020. PMID: 32477409 Free PMC article.
-
Global expression profiling identifies a novel biosignature for protein aggregation R120GCryAB cardiomyopathy in mice.Physiol Genomics. 2008 Oct 8;35(2):165-72. doi: 10.1152/physiolgenomics.00297.2007. Epub 2008 Jul 15. Physiol Genomics. 2008. PMID: 18628338 Free PMC article.
-
Metabolic Reprogramming in Modulating T Cell Reactive Oxygen Species Generation and Antioxidant Capacity.Front Immunol. 2018 May 16;9:1075. doi: 10.3389/fimmu.2018.01075. eCollection 2018. Front Immunol. 2018. PMID: 29868027 Free PMC article. Review.
References
-
- Aouida, M., Page, N., Leduc, A., Peter, M., and Ramotar, D. (2004). A genome-wide screen in Saccharomyces cerevisiae reveals altered transport as a mechanism of resistance to the anticancer drug bleomycin. Cancer Res. 64, 1102-1109. - PubMed
-
- Balshaw, D. M., Xu, L., Yamaguchi, N., Pasek, D. A., and Meissner, G. (2001). Calmodulin binding and inhibition of cardiac muscle calcium release channel (ryanodine receptor). J. Biol. Chem. 276, 20144-20153. - PubMed
-
- Bannister, S. J., and Wittrup, K. D. (2000). Glutathione excretion in response to heterologous protein secretion in Saccharomyces cerevisiae. Biotechnol. Bioeng. 68, 389-395. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
