

Monitoring neural activity and [Ca2+] with genetically encoded Ca2+ indicators
- PMID: 15509744
- PMCID: PMC6730159
- DOI: 10.1523/JNEUROSCI.2854-04.2004
Monitoring neural activity and [Ca2+] with genetically encoded Ca2+ indicators
Abstract
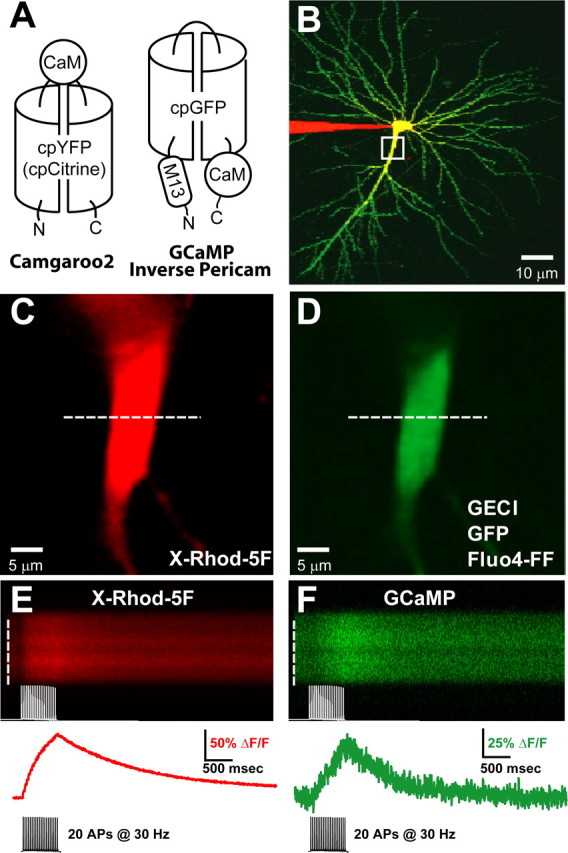
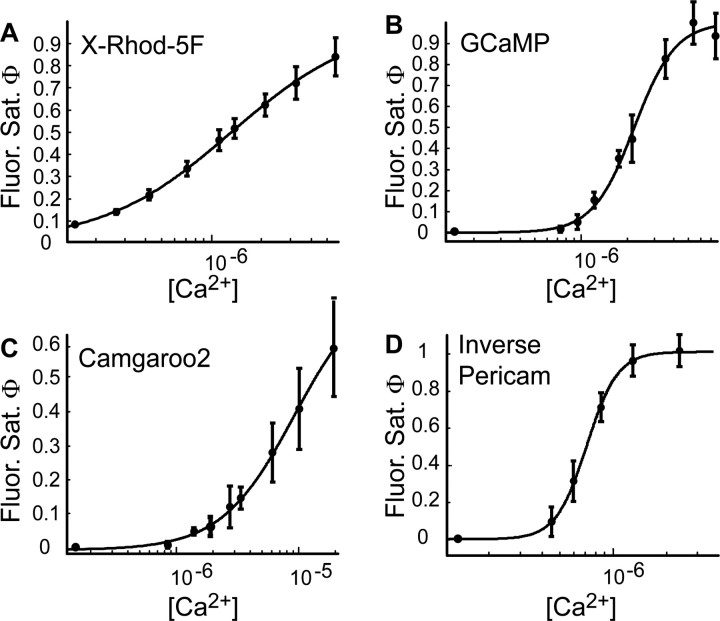
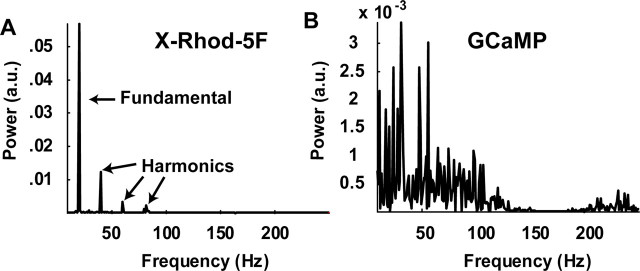
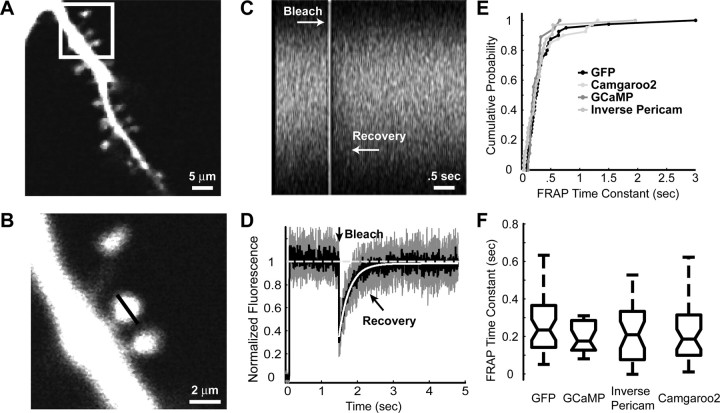
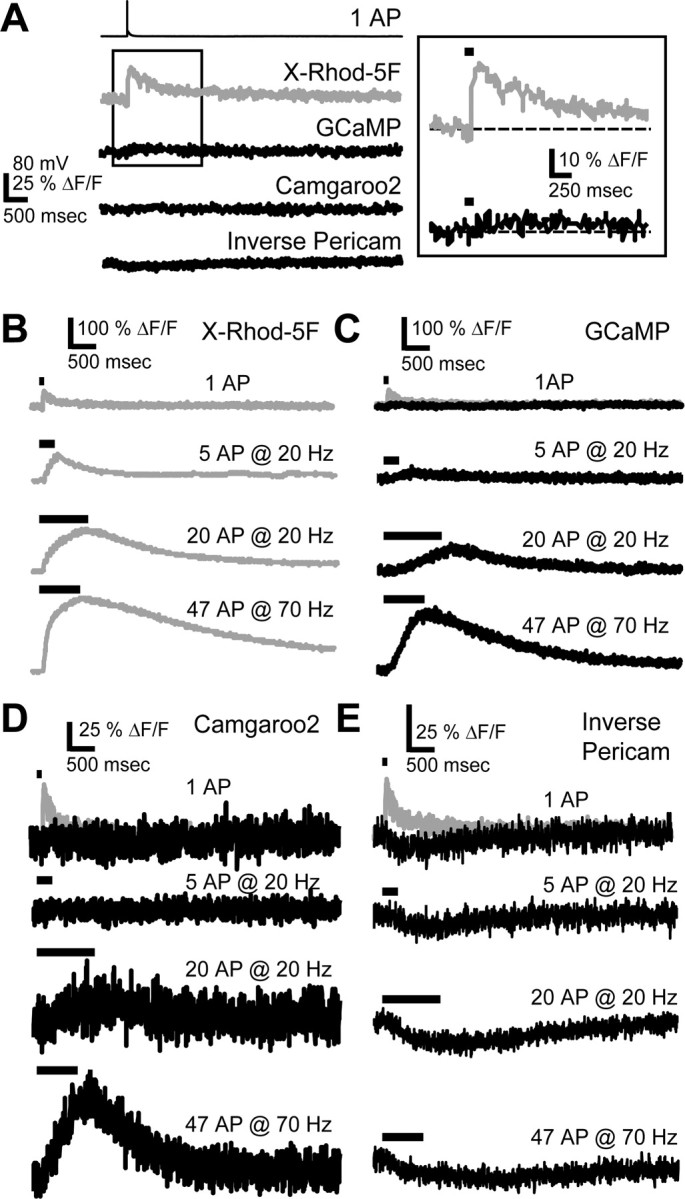
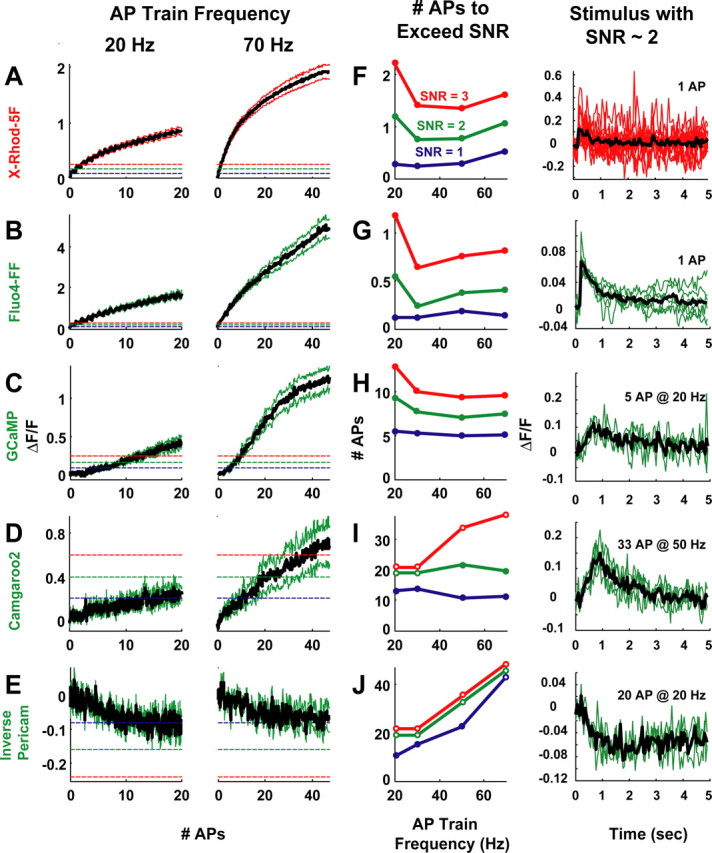
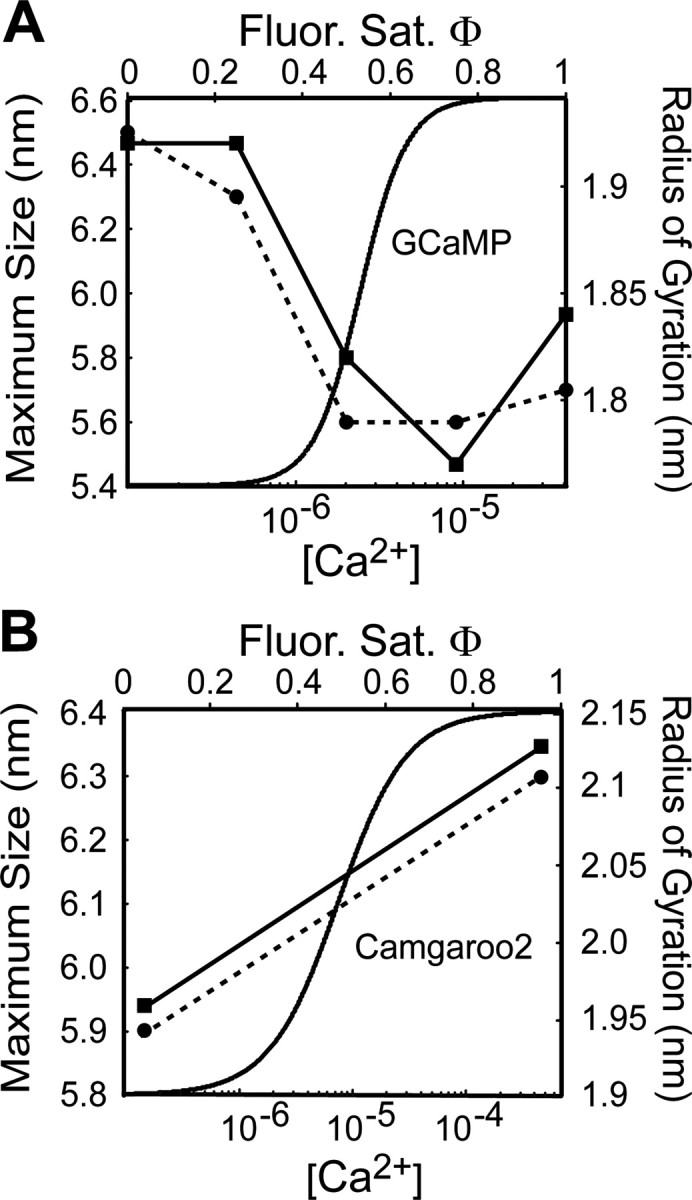
Genetically encoded Ca2+ indicators (GECIs) based on fluorescent proteins (XFPs) and Ca2+-binding proteins [like calmodulin (CaM)] have great potential for the study of subcellular Ca2+ signaling and for monitoring activity in populations of neurons. However, interpreting GECI fluorescence in terms of neural activity and cytoplasmic-free Ca2+ concentration ([Ca2+]) is complicated by the nonlinear interactions between Ca2+ binding and GECI fluorescence. We have characterized GECIs in pyramidal neurons in cultured hippocampal brain slices, focusing on indicators based on circularly permuted XFPs [GCaMP (Nakai et al., 2001), Camgaroo2 (Griesbeck et al., 2001), and Inverse Pericam (Nagai et al., 2001)]. Measurements of fluorescence changes evoked by trains of action potentials revealed that GECIs have little sensitivity at low action potential frequencies compared with synthetic [Ca2+] indicators with similar affinities for Ca2+. The sensitivity of GECIs improved for high-frequency trains of action potentials, indicating that GECIs are supralinear indicators of neural activity. Simultaneous measurement of GECI fluorescence and [Ca2+] revealed supralinear relationships. We compared GECI fluorescence saturation with CaM Ca2+-dependent structural transitions. Our data suggest that GCaMP and Camgaroo2 report CaM structural transitions in the presence and absence of CaM-binding peptide, respectively.
Figures
References
-
- Berridge MJ (1998) Neuronal calcium signaling. Neuron 21: 13-26. - PubMed
-
- Callaway JC, Ross WN (1995) Frequency-dependent propagation of sodium action potentials in dendrites of hippocampal CA1 pyramidal neurons. J Neurophysiol 74: 1395-1403. - PubMed
-
- Demaurex N, Frieden M (2003) Measurements of the free luminal ER Ca(2+) concentration with targeted “cameleon” fluorescent proteins. Cell Calcium 34: 109-119. - PubMed
-
- Evenas J, Malmendal A, Thulin E, Carlstrom G, Forsen S (1998) Ca2+ binding and conformational changes in a calmodulin domain. Biochemistry 37: 13744-13754. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous