

Biochemical interactions between proteins and mat1 cis-acting sequences required for imprinting in fission yeast
- PMID: 15509785
- PMCID: PMC525487
- DOI: 10.1128/MCB.24.22.9813-9822.2004
Biochemical interactions between proteins and mat1 cis-acting sequences required for imprinting in fission yeast
Abstract
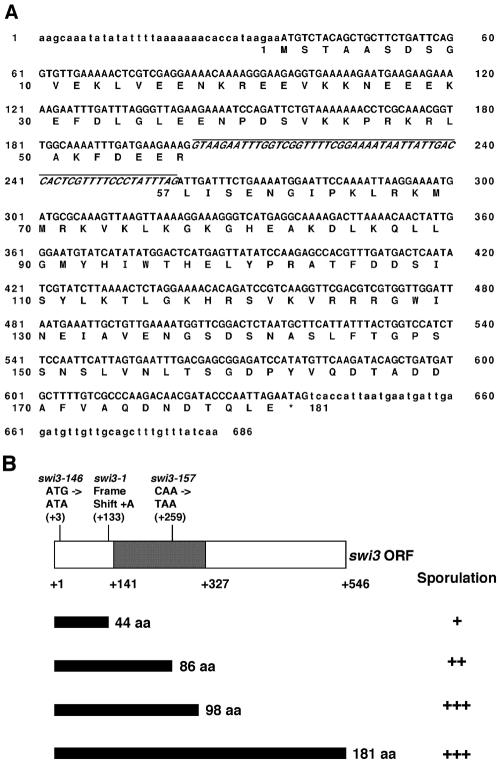
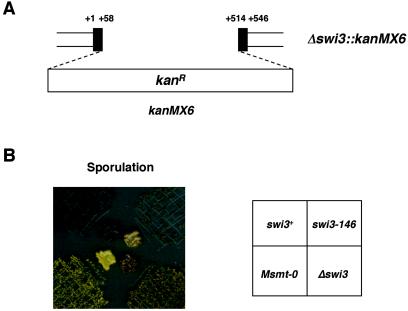
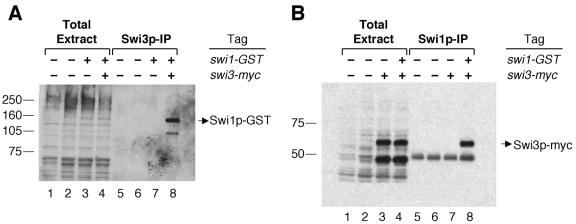
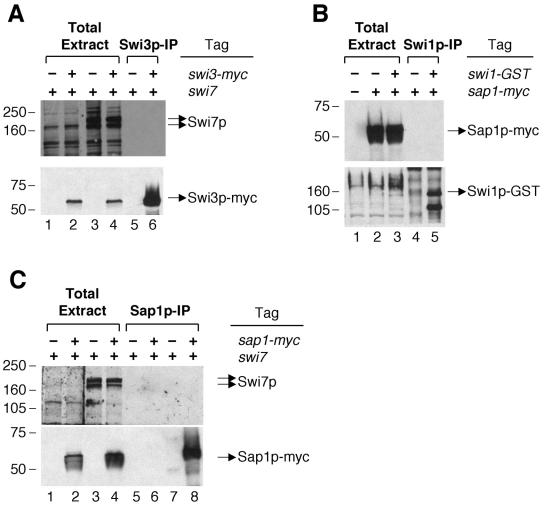
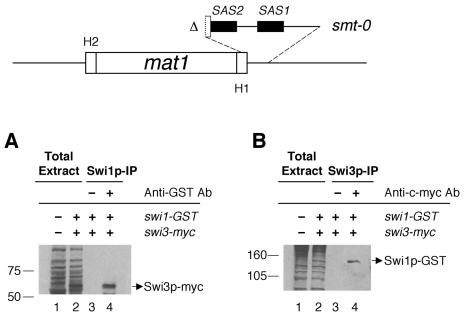
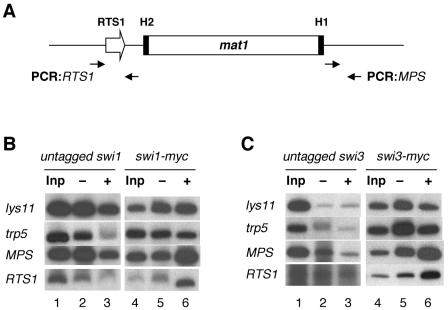
DNA recombination required for mating type (mat1) switching in Schizosaccharomyces pombe is initiated by mat1 imprinting. The imprinting event is regulated by mat1 cis-acting elements and by several trans-acting factors, including swi1 (for switch), swi3, swi7, and sap1. swi1 and swi3 were previously shown to function in dictating unidirectional mat1 DNA replication by controlling replication fork movement around the mat1 region and, second, by pausing fork progression around the imprint site. With biochemical studies, we investigated whether the trans-acting factors function indirectly or directly by binding to the mat1 cis-acting sequences. First, we report the identification and DNA sequence of the swi3 gene. swi3 is not essential for viability, and, like the other factors, it exerts a stimulatory effect on imprinting. Second, we showed that only Swi1p and Swi3p interact to form a multiprotein complex and that complex formation did not require their binding to a DNA region defined by the smt-0 mutation. Third, we found that the Swi1p-Swi3p complex physically binds to a region around the imprint site where pausing of replication occurs. Fourth, the protein complex also interacted with the mat1-proximal polar terminator of replication (RTS1). These results suggest that the stimulatory effect of swi1 and swi3 on switching and imprinting occurs through interaction of the Swi1p-Swi3p complex with the mat1 regions.
Figures
Similar articles
-
swi1 and swi3 perform imprinting, pausing, and termination of DNA replication in S. pombe.Cell. 2000 Sep 15;102(6):745-51. doi: 10.1016/s0092-8674(00)00063-5. Cell. 2000. PMID: 11030618
-
Fission yeast Swi1-Swi3 complex facilitates DNA binding of Mrc1.J Biol Chem. 2010 Dec 17;285(51):39609-22. doi: 10.1074/jbc.M110.173344. Epub 2010 Oct 5. J Biol Chem. 2010. PMID: 20924116 Free PMC article.
-
RNase-sensitive DNA modification(s) initiates S. pombe mating-type switching.Genes Dev. 2004 Apr 1;18(7):794-804. doi: 10.1101/gad.289404. Epub 2004 Apr 1. Genes Dev. 2004. PMID: 15059961 Free PMC article.
-
RTS1-an eukaryotic terminator of replication.Int J Biochem Cell Biol. 2002 Sep;34(9):1031-4. doi: 10.1016/s1357-2725(02)00040-7. Int J Biochem Cell Biol. 2002. PMID: 12009298 Review.
-
RNA insertion in DNA as the imprint moiety: the fission yeast paradigm.Curr Genet. 2019 Dec;65(6):1301-1306. doi: 10.1007/s00294-019-00991-x. Epub 2019 May 10. Curr Genet. 2019. PMID: 31076844 Review.
Cited by
-
Swi1 associates with chromatin through the DDT domain and recruits Swi3 to preserve genomic integrity.PLoS One. 2012;7(8):e43988. doi: 10.1371/journal.pone.0043988. Epub 2012 Aug 30. PLoS One. 2012. PMID: 22952839 Free PMC article.
-
The mating type switch-activating protein Sap1 Is required for replication fork arrest at the rRNA genes of fission yeast.Mol Cell Biol. 2005 Oct;25(19):8755-61. doi: 10.1128/MCB.25.19.8755-8761.2005. Mol Cell Biol. 2005. PMID: 16166653 Free PMC article.
-
Schizosaccharomyces pombe Swi1, Swi3, and Hsk1 are components of a novel S-phase response pathway to alkylation damage.Mol Cell Biol. 2005 Apr;25(7):2770-84. doi: 10.1128/MCB.25.7.2770-2784.2005. Mol Cell Biol. 2005. PMID: 15767681 Free PMC article.
-
Mammalian TIMELESS and Tipin are evolutionarily conserved replication fork-associated factors.J Mol Biol. 2007 Feb 9;366(1):36-52. doi: 10.1016/j.jmb.2006.10.097. Epub 2006 Nov 3. J Mol Biol. 2007. PMID: 17141802 Free PMC article.
-
Swi1Timeless Prevents Repeat Instability at Fission Yeast Telomeres.PLoS Genet. 2016 Mar 18;12(3):e1005943. doi: 10.1371/journal.pgen.1005943. eCollection 2016 Mar. PLoS Genet. 2016. PMID: 26990647 Free PMC article.
References
-
- Ahmed, S., S. Saini, S. Arora, and J. Singh. 2001. Chromodomain protein Swi6-mediated role of DNA polymerase alpha in establishment of silencing in fission yeast. J. Biol. Chem. 276:47814-47821. - PubMed
-
- Bahler, J., J.-Q. Wu, M. S. Longtine, N. G. Shah, A. McKenzie III, A. B. Steever, A. Wach, P. Philippsen, and J. R. Pringle. 1998. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14:943-951. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases