

Three myosin V structures delineate essential features of chemo-mechanical transduction
- PMID: 15510214
- PMCID: PMC533045
- DOI: 10.1038/sj.emboj.7600458
Three myosin V structures delineate essential features of chemo-mechanical transduction
Abstract
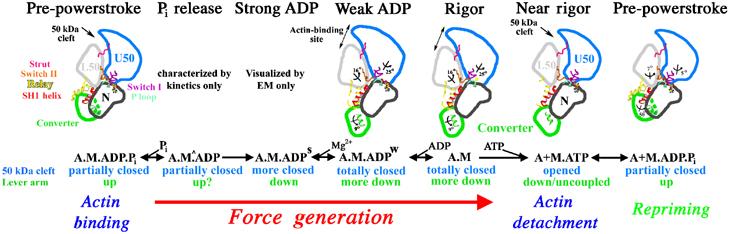
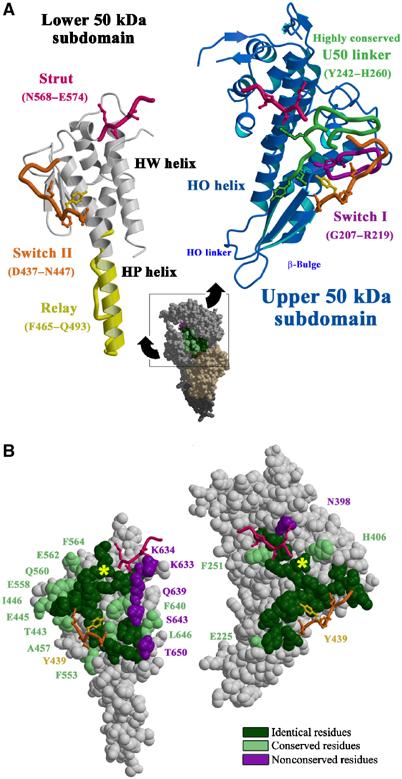
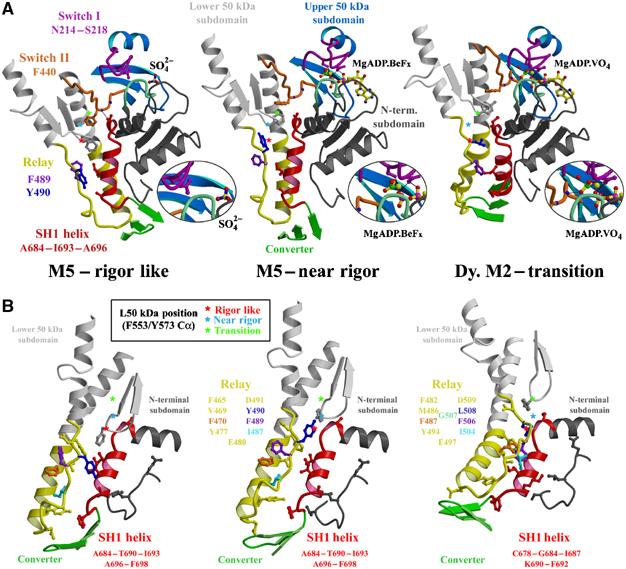
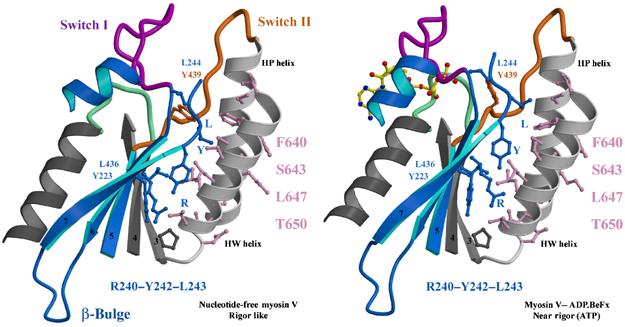
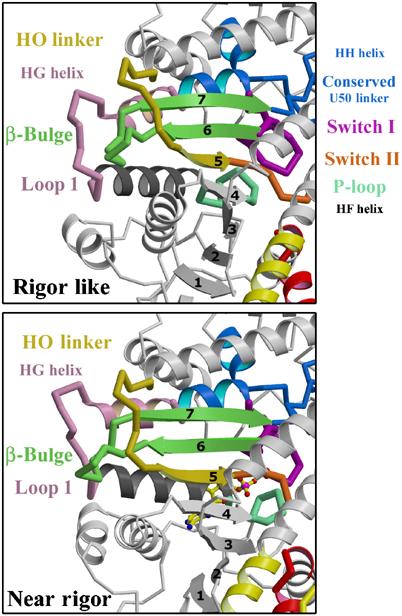
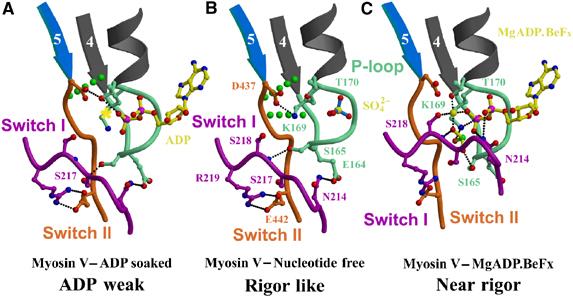
The molecular motor, myosin, undergoes conformational changes in order to convert chemical energy into force production. Based on kinetic and structural considerations, we assert that three crystal forms of the myosin V motor delineate the conformational changes that myosin motors undergo upon detachment from actin. First, a motor domain structure demonstrates that nucleotide-free myosin V adopts a specific state (rigor-like) that is not influenced by crystal packing. A second structure reveals an actomyosin state that favors rapid release of ADP, and differs from the rigor-like state by a P-loop rearrangement. Comparison of these structures with a third structure, a 2.0 angstroms resolution structure of the motor bound to an ATP analog, illuminates the structural features that provide communication between the actin interface and nucleotide-binding site. Paramount among these is a region we name the transducer, which is composed of the seven-stranded beta-sheet and associated loops and linkers. Reminiscent of the beta-sheet distortion of the F1-ATPase, sequential distortion of this transducer region likely controls sequential release of products from the nucleotide pocket during force generation.
Figures

References
-
- Bernstein SI, Milligan RA (1997) Fine tuning a molecular motor: the location of alternative domains in the Drosophila myosin head. J Mol Biol 271: 1–6 - PubMed
-
- Collaborative Computational Project No. 4 (1994) The CCP4 suite: programs for protein crystallography. Acta Crystallogr D 50: 760–763 - PubMed
-
- Coureux P-D, Wells AL, Menetrey J, Yengo CM, Morris CA, Sweeney HL, Houdusse A (2003) A structural state of the myosin V motor without bound nucleotide. Nature 425: 419–423 - PubMed
-
- Fisher AJ, Smith CA, Thoden JB, Smith R, Sutoh K, Holden HM, Rayment I (1995) X-ray structures of the myosin motor domain of Dictyostelium discoideum complexed with MgADP.BeFx and MgADP.AlF4−. Biochemistry 34: 8960–8972 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
