

A mitochondrial pool of sphingomyelin is involved in TNFalpha-induced Bax translocation to mitochondria
- PMID: 15516208
- PMCID: PMC1134862
- DOI: 10.1042/BJ20041627
A mitochondrial pool of sphingomyelin is involved in TNFalpha-induced Bax translocation to mitochondria
Abstract
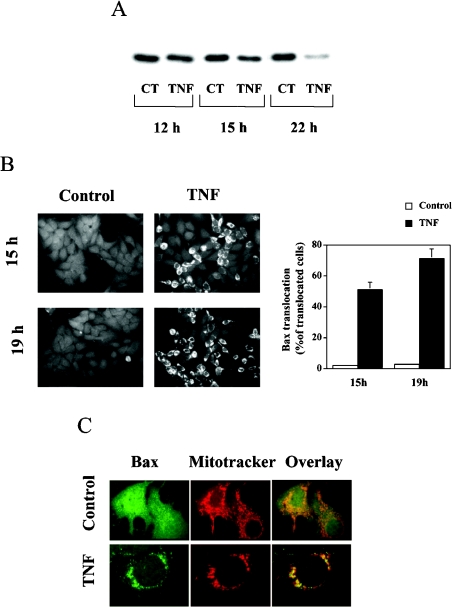
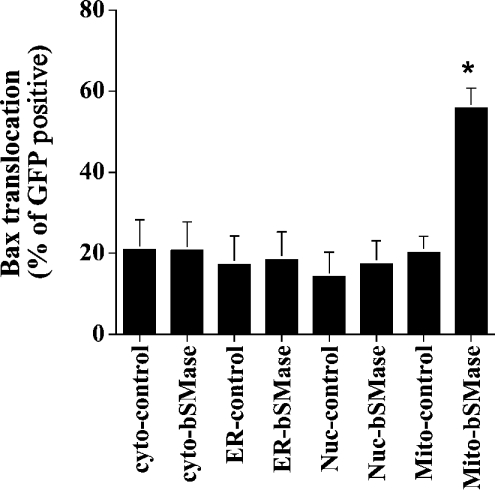
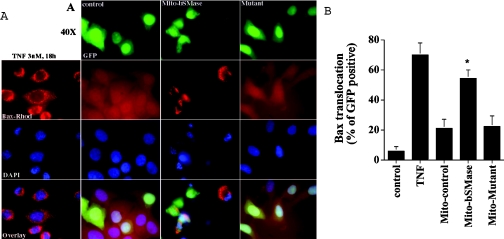
We recently showed that targeting bSMase (bacterial sphingomyelinase) specifically to mitochondria caused accumulation of ceramide in mitochondria, and induced cytochrome c release and cell death [Birbes, El Bawab, Hannun and Obeid (2001) FASEB J., 15, 2669-2679]. In the present study, we investigated the role of this mitochondrial pool of ceramide in response to a receptor-mediated event, namely TNFalpha (tumour necrosis factor alpha), and the involvement of this mitochondrial pool of ceramide in Bax translocation to mitochondria, an event that precedes cytochrome c release. Treatment of MCF7 cells with TNFalpha caused an increase in ceramide levels in the mitochondrial fraction which accompanied Bax translocation to mitochondria. Targeting bSMase to mitochondria specifically resulted in Bax translocation to mitochondria, suggesting that the mitochondrial ceramide pool is involved in Bax translocation. Moreover, in a reconstituted cell-free system, treatment of isolated mitochondria with bSMase enhanced Bax association with mitochondrial membranes. Collectively, these results suggest that the generation of ceramide in mitochondria in response to TNFalpha is sufficient to induce Bax translocation to mitochondria and subsequent cytochrome c release and cell death.
Figures
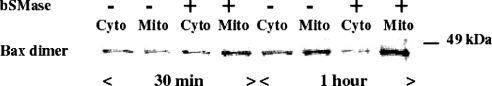
References
-
- Li P., Nijhawan D., Budihardjo I., Srinivasula S. M., Ahmad M., Alnemri E. S., Wang X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell. 1997;91:479–489. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials