

Characteristics of zinc transport by two bacterial cation diffusion facilitators from Ralstonia metallidurans CH34 and Escherichia coli
- PMID: 15516561
- PMCID: PMC524879
- DOI: 10.1128/JB.186.22.7499-7507.2004
Characteristics of zinc transport by two bacterial cation diffusion facilitators from Ralstonia metallidurans CH34 and Escherichia coli
Abstract
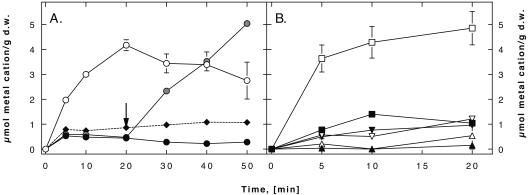
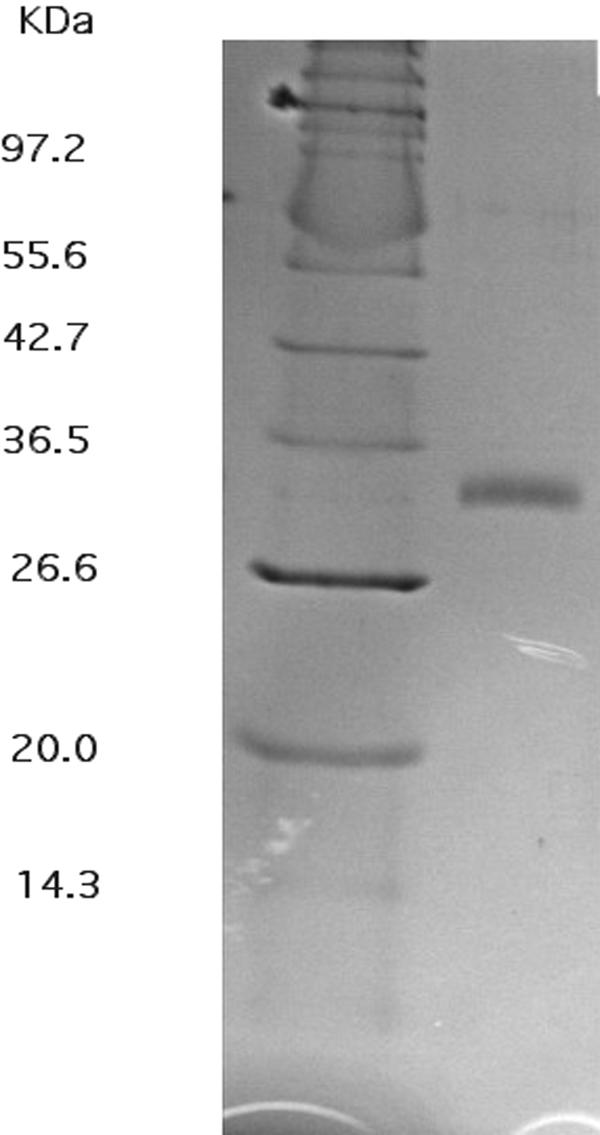
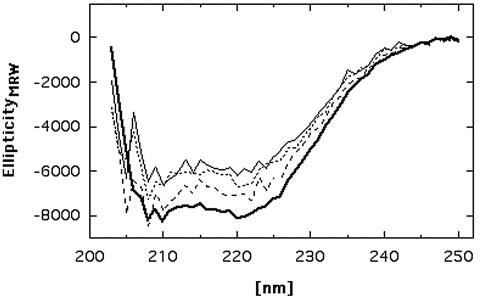
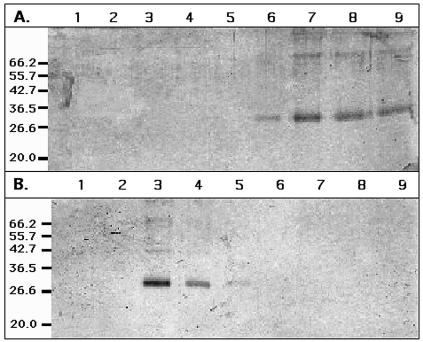
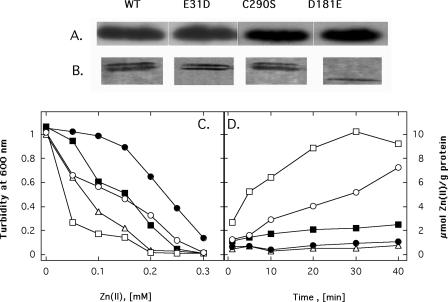
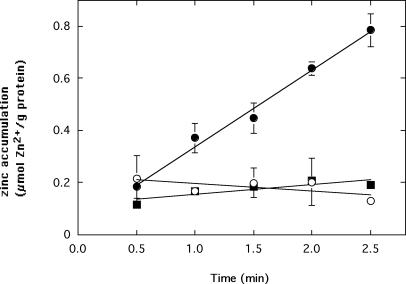
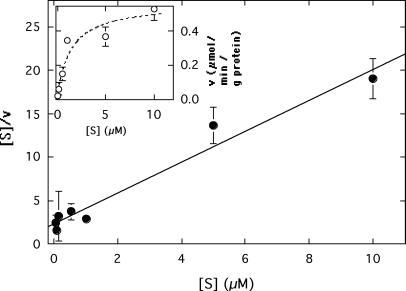
CzcD from Ralstonia metallidurans and ZitB from Escherichia coli are prototypes of bacterial members of the cation diffusion facilitator (CDF) protein family. Expression of the czcD gene in an E. coli mutant strain devoid of zitB and the gene for the zinc-transporting P-type ATPase zntA rendered this strain more zinc resistant and caused decreased accumulation of zinc. CzcD, purified as an amino-terminal streptavidin-tagged protein, bound Zn2+, Co2+, Cu2+, and Ni2+ but not Mg2+, Mn2+, or Cd2+, as shown by metal affinity chromatography. Histidine residues were involved in the binding of 2 to 3 mol of Zn2+ per mol of CzcD. ZitB transported 65Zn2+ in the presence of NADH into everted membrane vesicles with an apparent Km of 1.4 microM and a Vmax of 0.57 nmol of Zn2+ min(-1) mg of protein(-1). Conserved amino acyl residues that might be involved in binding and transport of zinc were mutated in CzcD and/or ZitB, and the influence on Zn2+ resistance was studied. Charged or polar amino acyl residues that were located within or adjacent to membrane-spanning regions of the proteins were essential for the full function of the proteins. Probably, these amino acyl residues constituted a pathway required for export of the heavy metal cations or for import of counter-flowing protons.
Figures
References
-
- Chao, Y., and D. Fu. 2004. Kinetic study of the antiport mechanism of an Escherichia coli zinc transporter, ZitB. J. Biol. Chem. 279:12043-12050. - PubMed
-
- Gaither, L. A., and D. J. Eide. 2001. Eukaryotic zinc transporters and their regulation. Biometals 14:251-270. - PubMed
-
- Goldberg, M., T. Pribyl, S. Juhnke, and D. H. Nies. 1999. Energetics and topology of CzcA, a cation/proton antiporter of the RND protein family. J. Biol. Chem. 274:26065-26070. - PubMed
-
- Goris, J., P. De Vos, T. Coenye, B. Hoste, D. Janssens, H. Brim, L. Diels, M. Mergeay, K. Kersters, and P. Vandamme. 2001. Classification of metal-resistant bacteria from industrial biotopes as Ralstonia campinensis sp. nov., Ralstonia metallidurans sp. nov. and Ralstonia basilensis Steinle et al. 1998 emend. Int. J. Syst. Evol. Microbiol. 51:1773-1782. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
