

Continuous control in bacterial regulatory circuits
- PMID: 15516575
- PMCID: PMC524909
- DOI: 10.1128/JB.186.22.7618-7625.2004
Continuous control in bacterial regulatory circuits
Abstract
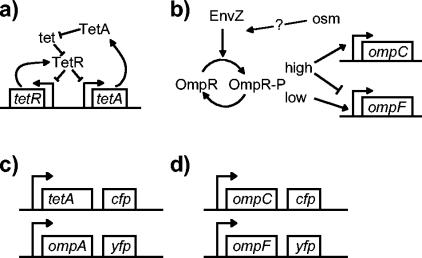
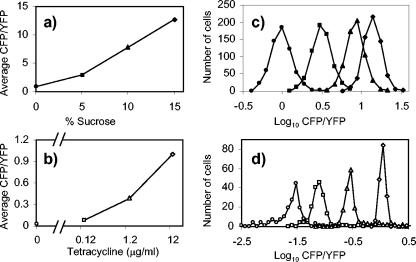
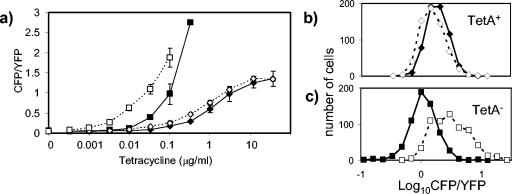
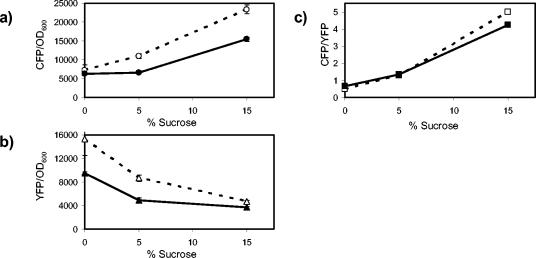
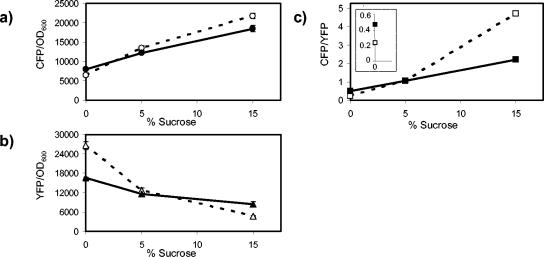
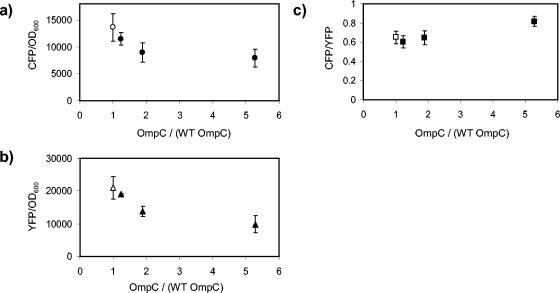
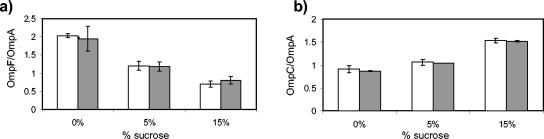
We show that for two well-characterized regulatory circuits in Escherichia coli, Tn10 tetracycline resistance and porin osmoregulation, the transcriptional outputs in individual cells are graded functions of the applied stimuli. These systems are therefore examples of naturally occurring regulatory circuits that exhibit continuous control of transcription. Surprisingly, however, we find that porin osmoregulation is open loop; i.e., the porin expression level does not feed back into the regulatory circuit. This mode of control is particularly interesting for an organism such as E. coli, which proliferates in diverse environments, and raises important questions regarding the biologically relevant inputs and outputs for this system.
Figures
References
-
- Amann, E., B. Ochs, and K. J. Abel. 1988. Tightly regulated tac promoter vectors useful for the expression of unfused and fused proteins in Escherichia coli. Gene 69:301-315. - PubMed
-
- Bachman, B. J. 1996. Derivations and genotypes of some mutant derivatives of Escherichia coli K-12, p. 2460-2488. In F. C. Neidhardt, R. Curtis, J. L. Ingraham, E. C. C. Lin, K. B. Low, B. Magasanik, W. S. Reznikoff, M. Riley, M. Schaecter, and H. E. Umbarger (ed.), Escherichia coli and Salmonella: cellular and molecular biology, 2nd ed., vol. 2. ASM Press, Washington, D.C.
-
- Bagowski, C. P., and J. E. Ferrell, Jr. 2001. Bistability in the JNK cascade. Curr. Biol. 11:1176-1182. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
